§ 7. Анализ некоторых частных примеров и их интерпретация

(Очевидно, что k < 1, а поскольку величина 1 — k представляет собой ту долю биомассы, которая расходуется на дыхание, обмен и т.
п., то в большинстве реальных экосистем k достаточно мало. Поэтому вместо формул (7.1) и (7.2)1 можно пользоваться более простыми приближениями, полученными из этих соотношений в предположении малости k.
' Если считать, что am ~ О (1), то оценки Q* (q) для незамкнутой и замкнутой цепей, сделанные с точностью до

Чтобы сравнить между собой характер роста длины трофической цепи при возрастании скорости поступления ре-
сурса для различных значений k, на рис. 30 приведены графики зависимостей q от безразмерной скорости поступления ресурса _ Кроме того, на рис. 31 при фиксиро
_ Кроме того, на рис. 31 при фиксиро
ванном значении Q показана длина цепи для отдельных значений k.
Для того чтобы проиллюстрировать, каким образом влияег на длину трофической цепи ее замыкание, и зависимость длины цепи q от степени замыкания а, на рис. 32 приведены графики зависимости критических скоростей
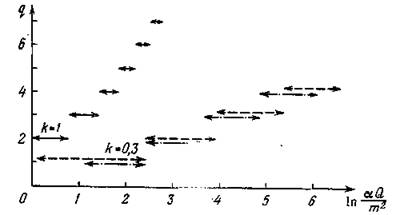
Рис. 30. Зависимость длины трофической цепи от скорости поступления внешнего ресурса при различных значениях k (в полулогарифмическом масштабе):
Z,Z. Z.Z расчет по приолижен-
Z.Z расчет по приолижен-
ной формуле (7.5).
Ф* (q) от а для нескольких значений q (при k — 1), которые выделяют в плоскости (Q, а) области существования цепей длины q — 1, 2, ...
Интерпретация всех этих зависимостей достаточно очевидна: длина трофической цепи увеличивается при возрастании k — к. п. д. перехода с одного трофического уровня на другой, — при возрастании а, которое можно интерпретировать либо как энергетическую ценность биомассы потребляемого вида, либо как ее доступность для потребителя, и при возрастании Q — скорости поступления в экосистему внешнего ресурса. При возрастании т — коэффициентов смертности — длина трофической цепи соответственно уменьшается. Замыкание трофической цепи приводит к тому, что при определенной степени замыкания длина цепи тоже увеличивается, но не более, чем на единицу.
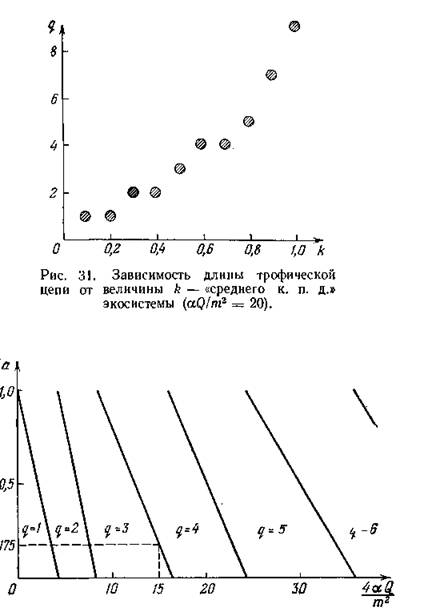
Рис. 32. Зависимость критических скоростей поступления ресурса 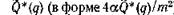 ) от степени замыкания цепи а. Если, например, при 4aQ/m2 = 15 длина незамкнутой трофической цепи равна 3, то при той же скорости поступления внешнего ресурса длина замкнутой цепи
) от степени замыкания цепи а. Если, например, при 4aQ/m2 = 15 длина незамкнутой трофической цепи равна 3, то при той же скорости поступления внешнего ресурса длина замкнутой цепи
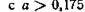 равна 4.
равна 4.
Характерно, что чем длиннее трофическая цепь, тем больший квант ресурса необходимо добавить в систему для закрепления в ней нового вида. В самом деле, как это следует из (5.6) и (6.9), для увеличения длины трофической цепи с q до q + 1 (закрепление в системе нового вида) скорость поступления ресурса должна увеличиться для незамкнутой цепи на Be.’””"”"’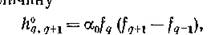
а для замкнутой цепи — на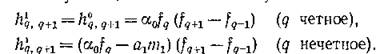 величину
величину
Легко видеть, что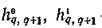 — монотонно возрастающие
— монотонно возрастающие
функции q. Например, при малых kt — k справедлива оценка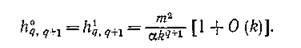
Из всего вышеизложенного следует, что чем длиннее трофическая цепь, тем с большим трудом происходит закрепление в системе нового вида (или нового трофического уровня). Наиболее интересный вывод: существуют дискретные уровни потока внешнего ресурса в экосистему, которые определяют длину трофической цепи. Другими словами, поступающая в экосистему энергия «квантуется», причем чем больше номер возможного трофического уровня в экосистеме, тем больший «квант» энергии должен в нее поступить.