ПУТИ РЕГУЛИРОВАНИЯ УРОВНЯ СВОБОДНОРАДИКАЛЬНЫХ ПРОДУКТОВ И АКТИВНЫХ КИСЛОРОДНЫХ МЕТАБОЛИТОВ
Регулирование уровня активных кислородных метаболитов и свободнорадикальных продуктов ПОЛ осуществляется антиоксидантными системами, снижающими активность радикальных окислительных процессов.
В настоящее время под антиоксидантами понимают широкий класс соединений, ингибирующих окислительные процессы по одному или нескольким механизмам: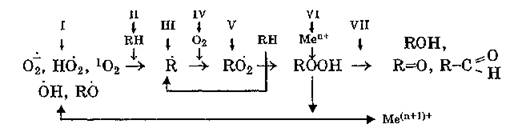
Различают превентивные антиоксиданты (пути: П — изменение структурной организации субстрата, замедляющее окисление; IV — снижение концентрации О2; VI — связывание или окисление ионов металлов переменной валентности, индуцирующих разложение пероксидов и образование радикалов; VII — перевод пероксидов в стабильные продукты окисления: спирты, альдегиды, кетоны) и ингибиторы АКМ (пути: I — ингибирование радикальных форм АКМ, способных инициировать образование органических радикалов; III и V — прерывание окислительной цепи посредством взаимодействия с органическими радикалами). Действие ингибиторов АКМ является специфичным.
Часто антиоксиданты классифицируют в соответствии с величинами их молекулярных масс. К группе высокомолекулярных соединений — ферментов антиоксидантной защиты и белков, связывающих катализаторы свободнорадикальных процессов — ионы Fe и Си, относят соответственно супероксиддисмута- зу (СОД), церулоплазмин, пероксидазу, каталазу, глутатионзави- симые ферменты и альбумин крови, трансферрин, ферритин. Группу низкомолекулярных антиоксидантов составляют а-токоферол, аскорбат, глутатион, мочевая кислота, мочевина, билирубин, некоторые аминокислоты.
Ферментативные антиоксиданты (АО) характеризуются высокой специфичностью действия, а также клеточной и органной локализации, использованием в качестве катализаторов некоторых металлов (Си, Zn, Fe, Se). Уровень внутриклеточных ферментативных АО находится под генетическим контролем.
У животных в условиях гипоксии и гипероксии, усиливающих образование АКМ, повышается уровень внутриклеточных ферментативных АО, что связано с механизмами поддержания устойчивости организмов к окислительному стрессу.Супероксиддисмутаза (КФ 1.15.1.11, СОД) катализирует реакцию дисмутации супероксидного анион-радикала: 2О,,'~ + 2Н+ -» Н2О2 + О2. Обнаружено несколько изоферментов этого белка, различающихся локализацией, строением активного центра и некоторыми физико-химическими свойствами. Си, Zn-содержащая СОД чувствительна к цианиду и содержится в цитозоле и в межмембранном пространстве клеток эукариот. Цианидрезистент- ная Мп-СОД (железосодержащий изофермент) локализована в митохондриях эукариот и найдена у прокариот. В плазме содержится цианидчувствительная экстрацеллюлярная СОД, представляющая собой Си, Zn-содержащую тетрамерную молекулу (Mr -120—135 кДа) из четырех гликопротеиновых субъединиц. Предполагают, что экстрацеллюлярная СОД выполняет функцию защиты клеток эндотелия во всем организме. Однако активность СОД в плазме крови намного ниже, чем для цитозольного фермента. По-видимому, это связано с накоплением конечного продукта реакции — пероксида водорода, являющегося ингибитором фермента. В клетках пероксид водорода быстро разрушается внутриклеточными каталазой и глутатионпероксидазой.
Несмотря на высокую специфичность фермента, в определенных условиях Cu-СОД может взаимодействовать с пероксидом водорода и выступать в качестве прооксиданта, инициируя образование радикалов 0,7 и ОН":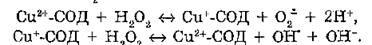
В настоящее время исследуются возможности клинического применения СОД, обладающей выраженным защитным эффектом при воспалительных, ишемических и стрессовых поражениях. С целью повышения стабильности фермента и предотвращения его быстрого разрушения во внеклеточном пространстве используют препараты фермента, ковалентно связанного с иммуноглобулинами, сывороточным альбумином, полиэтиленгликолем.
Каталаза (КФ 1.11.1.6) ускоряет процесс двухэлектронного восстановления пероксида водорода до воды, используя Н2О2 как донор электрона. Молекула каталазы состоит из четырех идентичных субъединиц и четырех групп гематина. Молекулярная масса фермента из различных источников составляет 225— 250 кДа. Каталаза локализована преимущественно в пероксисо- мах клеток, где ее концентрация достигает - 10“° моль/л. Максимальное содержание фермента обнаружено в эритроцитах, печени и почках. Разложение пероксида водорода каталазой осуществляется в два этапа:
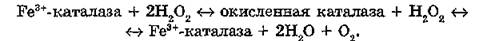
В окисленном состоянии каталаза работает и как пероксида- за, катализируя реакции окисления спиртов или альдегидов:
окисленная каталаза + АН2 —> 1Ге3+-каталаза + 2Н2О + А, где АН2 - донор электронов.
Церулоплазмин — один из основных антиоксидантов плазмы, проявляющий как специфическую, так и неспецифическую антиоксидантную активность. Специфическая активность, связанная со снижением уровня АКМ, обусловлена реализацией трех возможных механизмов: 1) церулоплазмин обладает феррокси- дазной активностью, окисляя ионы Fe2+ без образования супероксидного анион-радикала; 2) он способен вызывать дисмута- цию супероксидных радикалов, которая имеет не ферментативный, а стехиометрический характер (это обусловливает ингибирование церулоплазмином ПОЛ в липопротеинах); 3) церулощ лазмин инактивирует АФК, генерируемые миелопероксидазой, и тем самым защищает антипротеиназу от окислительного повреждения гипохлоритом.
Неспецифическая антиоксидантная активность церулоплазмина связана с образованием комплексных соединений с медью, что препятствует возможности их участия в реакциях Фентона и Габер—Вейса.
Антиоксидантной активностью обладает также трансферрин плазмы. Его действие носит в основном неспецифический характер, обусловленный связыванием ионов железа.
При насыщении железом белка до 100 % трансферрин может проявлять прооксидантное действие, связанное с генерацией гидроксильных радикалов. Избыток железа при полном насыщении трансферрина связывается неспецифически с поверхностью белка и в такой форме может участвовать в реакции Габер-Вейса.К группе высокомолекулярных антиоксидантов относятся глу- татионзависимые ферменты — глутатионпероксидаза, глутати- онредуктаза, глутатионтрансфераза.
Глутатионперокидаза (КФ 1.11.1.9) эффективно утилизирует токсичные липопероксиды в организме при помощи ферментативной реакции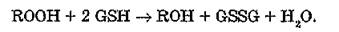
В тканях млекопитающих обнаружены по крайней мере два фермента, способные легко восстанавливать органические гидропероксиды. Селенсодержащая глутатионпероксидаза с высокой скоростью утилизирует как пероксид водорода, так и органические гидропероксиды, в том числе липопероксиды — пероксиды жирных кислот, ацилглицеридов, стероидов и простагландинов. Неселеновая глутатионпероксидаза II, напротив, активно восстанавливает исключительно органические гидропероксиды. Se-co- держащая глутатионпероксидаза локализована в цитозоле (-70 %) и митохондриях (20—30 %) клеток млекопитающих.
В регуляции обмена липопероксидов (рис. 29) важную роль играет сопряженное действие глутатионпероксидазы и глутати- онредуктазы (КФ 1,6.4.1), приводящее к инактивации пероксидов жирных кислот и превращению их в соответствующие оксикислоты. Глутатионредуктаза катализирует восстановление окисленного глутатиона:
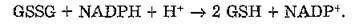
Глутатионтрансферазы (КФ 2.5.1.18) представляют собой группу ферментов, катализирующих начальную стадию биосинтеза меркаптуратов — конъюгацию глутатиона с ксенобиотиками (RX), содержащими электрофильный атом:
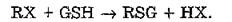
Однако продуктом ферментативного превращения некоторых субстратов, в том числе органических гидропероксидов, является не тиоэфир (RSG), а окисленный глутатион.
Образование GSSG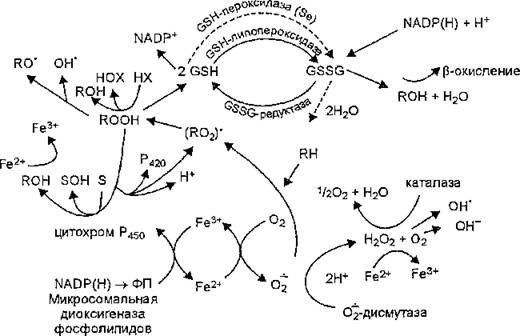
Рис. 29. Детоксикация супероксидного радикала, пероксида водорода и липопероксидов в тканях млекопитающих
при действии GSH-трансферазы на органические гидроперокси- ды объясняется взаимодействием электрофильного кислорода с тиолатным анионом (GS ), что приводит к возникновению нестабильного сульфенового производного глутатиона:
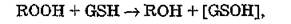
которое затем неферментативно реагирует с еще одной молекулой GSH:
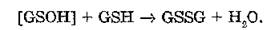
Глутатионтрансферазы локализованы преимущественно в цитозоле клеток; в печени человека они составляют 2—4 % от общего количества цитозольного белка. Глутатионтрансферазы эффективно восстанавливают гидрофобные гидропероксиды с большим объемом молекулы (гидропероксиды линолевой и ара- хидоновой полиненасыщенных жирных кислот, фосфолипидов), а также гидропероксиды мононуклеотидов и ДНК, участвуя тем самым в их репарации.
Считают, что при окислительном стрессе ферментативная антиоксидантная защита оказывается малоэффективной по сравнению с действием низкомолекулярных соединений. Это связано с тем, что ферменты-антиоксиданты локализованы внутри клеток, а в биологических жидкостях обнаруживаются лишь следовые их количества. Кроме того, при оксидативном повреждении происходит быстрая инактивация конститутивного пула ферментов свободными радикалами. Поэтому необходимо значительное время для индукции их синтеза. В этих условиях повышается роль низкомолекулярных антиоксидантов, не только снижающих интенсивность свободнорадикальных процессов, но и выполняющих важные метаболические функции.
Эффективными перехватчиками свободных радикалов являются фенольные антиоксиданты, содержащие ароматическое кольцо, связанное с одной или несколькими гидроксильными группами.
Благодаря наличию в структуре ароматического кольца обобщенной системы д-электронов происходит смещение отрицательного заряда на кислород, в результате которого осуществляется достаточно легкий отрыв атома водорода ОН-группы с образованием разных изомерных форм фенокси-радикала. Такие соединения выступают в качестве перехватчиков пероксидных и алкоксильных радикалов в следующих реакциях: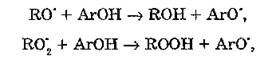
где АгОН — ароматическое кольцо фенольного антиоксиданта, связанное с гидроксильной группой.
Взаимодействие фенольных антиоксидантов с органическими радикалами приводит к образованию феноксильных радикалов (АгО‘), которые могут участвовать в реакциях диспропорционирования с образованием хинолидных пероксидов:
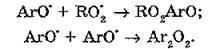
Распад хинолидных пероксидов приводит к образованию хинонных форм молекул, которые в нормальных условиях не обладают антиоксидантными свойствами, однако при дефиците кислорода могут тормозить окисление путем взаимодействия с алкильными радикалами:
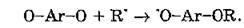
К фенольным соединениям, обладающим выраженной антиоксидантной активностью, относят витамины Б (а-токоферол) и К, триптофан, тирозин, фенилаланин, убихиноны, каротиноиды, флавоноиды, фенокарбоксильные кислоты и др. Они ингибируют супероксидный анион-радикал кислорода, синглетный молекулярный кислород, гидроксильный радикал и индуцированные ими процессы ПОЛ.
Вместе с тем благодаря способности легко отдавать и захватывать электроны фенольные АО могут выступать и в качестве восстановителей. Например, в условиях гипоксии при действии ряда дыхательных ядов в митохондриях убихинон окисляется кислородом с образованием супероксидного анион-радикала, т. е. проявляет прооксидантные свойства. а-Токоферол способен восстанавливать ионы металлов переменной валентности и действовать как прооксидант, в частности, при индуцированном ионами железа окислении липосом. Взаимодействие фенольных АО с пероксидами приводит к образованию алкоксильных радикалов, которые могут индуцировать окислительные реакции:
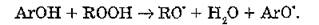
Аскорбиновая кислота способна выступать в качестве донора и акцептора водорода благодаря наличию в ее структуре двух енольных групп. Аскорбат образуется у животных в виде конечного продукта из разветвлений глюкуронатного пути обмена глюкозы. При окислении витамина С в тканях животных и человека образуется дегидроаскорбат, превращающийся затем в дике- тогулонат. При расщеплении последнего образуется щавелевая кислота, а при декарбоксилировании — ксилулоза, превращающаяся в глюкозу. У растений окисление аскорбата с образованием дегидроаскорбата катализирует медьсодержащая аскорбат- оксидаза. Эту же функцию выполняют и другие терминальные оксидазы растительных и животных тканей — цитохромоксида- за, фенолоксидаза, некоторые металлы. Одновременно с окисле- ни-ем аскорбата в организме происходит ферментативное и неферментативное восстановление дегидроаскорбата до аскорбата, осуществляющееся при участии восстановленного глутатиона. Второй путь быстрого образования аскорбата в организме — восстановление монодегидроаскорбата при участии NADH.
Способность аскорбата и дегидроаскорбата легко подвергаться окислительно-восстановительным превращениям лежит в основе широкого участия витамина С в обмене веществ: дыхании и фотосинтетической активности, транспорте электронов, окислении и восстановлении никотинамидных коферментов.
Аскорбат необходим для нормального развития организма, повышает эффективность адаптации к воздействию факторов внешней среды, участвует в обмене белков, липидов, углеводов, ускоряет процессы регенерации, стимулирует деятельность желез внутренней секреции.
Окисленная и восстановленная формы аскорбата способны проникать через мембранные структуры животных клеток. Благодаря большей липофильности дегидроаскорбат легко диффундирует через биомембраны, чем существенно отличается от аскорбата. Дегидроаскорбат как транспортная форма витамина С проникает в биологические структуры посредством простой или облегченной диффузии. В нормальных физиологических условиях основную роль в процессах трансмембранного переноса и поддержания физиологического уровня аскорбата в клетках играет механизм активного транспорта, опосредованный участием специфических белков и зависящий от наличия энергии.
Аскорбиновая кислота обладает чрезвычайно широким спектром антиоксидантных свойств: обезвреживает гипогалоиды, супероксидный анион-радикал кислорода, пергидроксильный радикал, пероксидный радикал, синглетный кислород, гидроксильный радикал, восстанавливает а-токоферильный радикал, возвращая а-токоферолу антиоксидантные свойства, а также тиильный (CS') и тиопероксильный (GSO‘a) радикалы. Витамин С превосходит другие антиоксиданты плазмы крови в защите липидов от пероксидного окисления, так как только это соединение достаточно реакционноспособно для эффективного ингибирования ПОЛ. Антиоксидантные свойства аскорбата основаны на функционировании одноэлектронных циклических переходов между дигидро-, семидигид- ро- и дегидроаскорбатными формами. Скорость этих превращений зависит от присутствия металлов переменной валентности, других пар окислитель—восстановитель и величины pH. Способностью непосредственно взаимодействовать с АФК характеризуется восстановленная форма аскорбиновой кислоты.
Витамины С, A, D и F, при окислении и аутоокислении которых образуются промежуточные радикальные формы, могут выполнять роль инициаторов окисления и ускорять ПОЛ, увеличивая скорость зарождения цепей реакций в мембранах. Кроме того, в присутствии ионов железа и меди аскорбиновая кислота становится мощным прооксидантом, что указывает на необходимость in vivo надежной секвестрации свободных ионов металлов пере
менной валентности. Проявление аскорбатом анти- или проок- сидантных свойств зависит также от концентрации субстрата и условий протекания окислительных реакций. Такая ее функциональная пластичность необходима для осуществления в биосистемах механизма редокс-регуляции.
Антиоксидантные свойства глутатиона (GSH) — трипептида, образованного цистеином, глутаминовой кислотой и глицином, определяются как непосредственным взаимодействием с АФК и обменными реакциями с дисульфидными связями, так и функционированием глутатионзависимых ферментов (см. выше). Механизм защитного эффекта глутатиона связан с инактивацией высокореакционных кислородных и органических свободных радикалов вследствие переноса атомов водорода с молекулы тиола. Образующиеся активные частицы обладают меньшим повреждающим эффектом по сравнению с радикалом, у которого неспаренный электрон локализован на атоме углерода:
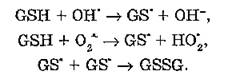
Тиильные радикалы эффективно удаляются из реакционной среды вследствие образования дисульфидов глутатиона.
Глутатион необходим для нормального функционирования эритроцитов и кроветворной ткани, в норме он практически весь ассоциирован с форменными элементами крови и находится в восстановленной форме (до 96 %). GSH участвует в восстановлении метгемоглобина. Ферменты синтеза глутатиона находятся в эритроцитах. Процесс восстановления его происходит с участием глутатионредуктазы, а также NADPH и зависит от активности пентозофосфатного пути. Клеточный глутатион участвует в поддержании пула восстановленного аскорбата: перенос восстановленных эквивалентов с GSH на аскорбат осуществляется глу- татиондегидроаскорбатредуктазой.
Мочевая кислота по своим свойствам близка к енольным антиоксидантам и может выступать синергистом с радикалами а-токоферола и аскорбиновой кислоты. Она способна инактивировать синглетный кислород, гидроксильный радикал, связывать ионы железа с участием аминогрупп, ингибировать ПОЛ и восстанавливать метгемоглобин с образованием радикала урата.
Разные группы антиоксидантов функционируют в тесной взаимосвязи: снижение концентрации или активности одних компонентов антиокислительных систем приводит к изменению уровня других, что обеспечивает поддержание окислительно-восстановительного гомеостаза клеток. Антиоксидантные ферменты и низкомолекулярные соединения катализируют последовательные этапы превращения АФК, например:
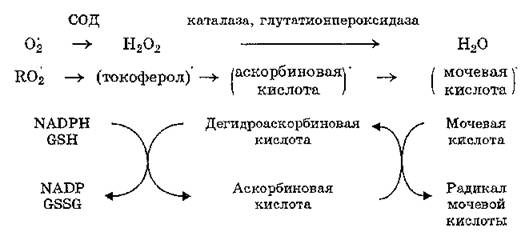
Высказано представление об антиоксидантных цепях переноса электронов, эффективность функционирования которых обеспечивается работой всех компонентов антиокислительных систем.
Антиоксиданты, модифицируя структуру и функциональную активность мембран, оказывают воздействие на клеточный метаболизм различными способами: в результате взаимодействия со свободными радикалами, рецепторами, путем ингибирования и активирования ферментов, непосредственного встраивания в мембрану, взаимодействия с генетическим аппаратом клетки (рис. 30).
Необходимо отметить, что выяснение множественных механизмов действия антиоксидантов в организме, включающих и воздействие их на скорость ПОЛ, и влияние на различные процессы клеточного метаболизма, представляет сложную научную задачу. Это связано с проявлением ими активности в широком интервале концентраций (10~18—10~2 моль/л) и реализацией в каждом интервале концентраций различных механизмов:
—• протеканием реакций ингибиторов со свободными радикалами;
— взаимодействием с клеточными рецепторами;
—• изменением текучести липидов мембран и мезофазных переходов в них;
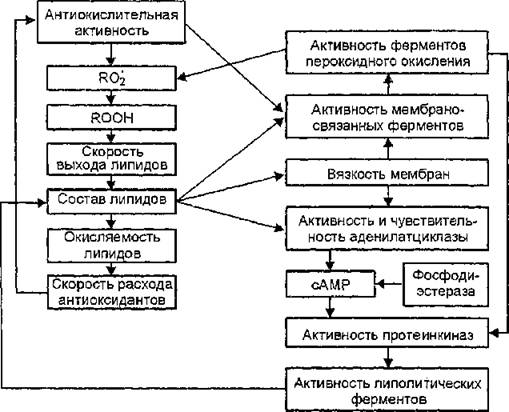
Рис. ЗО, Схема процессов регуляции клеточного метаболизма с участием окислительных реакций в липидах мембран (К. Е. Круглякова, Л. Н. Шишкина, 1992)
— непосредственным влиянием на активность ферментов;
— влиянием на лиганд-белковые отношения через параметрический резонанс.
Переход от одного механизма к другому обусловливает сложную немонотонную, полимодальную зависимость "доза—эффект" (Е. Б. Бурлакова, 1998).
КОНТРОЛЬНЫЕ ВОПРОСЫ
1. Опишите основные механизмы модификации белковых и липидных компонентов биомембран при воздействии экстремальных факторов внешней среды и развитии патологических состояний организма.
2. Что такое пероксидное окисление липидов, какие факторы инициируют этот процесс, какова последовательность стадий его развития?
3. В чем состоит сущность цепного окисления липидов?
4. В чем разница между ферментативным и неферментативным пероксидным окислением липидов?
5. Что такое активные формы кислорода, какова их роль в развитии различных патологических состояний организма?
6. Каковы механизмы образования различных активных форм кислорода?
7. Опишите свойства и пути утилизации супероксидного анион-радикала и пероксида водорода.
8. В чем отличие синглетного молекулярного кислорода от других активных кислородных метаболитов?
9. Какова роль NO-радикала в регулировании процессов клеточного метаболизма?
10. Что называют оксидативной модификацией макромолекул и окислительным стрессом?
11. Дайте определение понятия "антиоксиданты”.
12. Каковы основные механизмы ингибирования антиоксидантами окислительных процессов?
13. Охарактеризуйте основные компоненты системы высокомолекулярных антиоксидантов.
14. Какова роль глутатионзависимых ферментов в регулировании уровня свободнорадикальных продуктов ПОЛ и активных кислородных метаболитов?
15. В чем отличия прооксидантов от антиоксидантов?
16. Опишите свойства, механизмы действия и метаболические функции основных низкомолекулярных компонентов антиоксидантной системы.
17. Обоснуйте утверждение: "Биомембраны — непременные участники совокупности процессов возникновения и развития ряда патологических состояний организма человека".