Пути регулирования активности векторных ферментов биомембран
Одним из наиболее актуальных вопросов современной мембранологии является выяснение принципов и механизмов регуляции векторных ферментов биомембран (в том числе Nah, К+-АТФазы), выполняющих разнообразные “жизненно важные” функции не только для отдельных мембранных структур, но и для клетки в целом.
Полифункциональный характер Na+, К+-АТФа- зы (см. раздел 1.2.4), т.е. сочетание в ней метаболической, транспортной и рецепторной функций, определяет существование достаточно сложных механизмов ее регуляции. Кроме того, изучение механизмов функционирования и регулирования транспортных АТФаз на уровне отдельных клеток и субклеточных компонентов актуально не только в теоретическом, но и в практическом аспекте для оценки степени и характера нарушений этих механизмов при некоторых патологических состояниях, связанных с изменением ионного состава среды и накоплением активных форм кислорода (см. главу 3). Рассмотрим основные пути регулирования функциональной активности одного из ключевых векторных ферментов плазматических мембран клеток - Na+, К'- АТФазы.По современным представлениям, Na+, К+-АТФаза является типичным липидзависимым ферментом: для формирования его функционально-активной конформации необходимы кислые липиды. Показано, что мембранные гликолипиды, локализованные преимущественно в наружной половине бислоя, обеспечивают правильную ориентацию Na+, К ' -АТФазнсго комплекса относительно плоскости мембраны (т.е. они отвечают за проявление векторных свойств фермента). Вместе с тем остается неясным, какова специфическая роль мембранных фосфолипидов в обеспечении транспортной функции Na+, К'-АТФазы. По-видимому, функционирование центров связывания нуклеотидов и катионов на молекуле фермента не зависит от липидов, тогда как для осуществления конформационных перестроек в ферментном белке важна его связь с мембранными липидами.
Кривые, описывающие зависимость ферментативной активности от концентрации липида, имеют сигмоидную форму, что свидетельствует о кооперативном характере связывания липидов с белком. Однако абсолютная потребность фермента в тех или иных фосфолипидах (или их полярных головках) экспериментально не доказана. Так, опыты по ферментативному превращению одних фосфолипидов в другие (например, фосфатидилсерина в фосфатидилэтаноламин) в препаратах мембраносвязанной Na+, К'-АТФазы показали, что фермент может функционировать без отрицательно заряженных фосфолипидов, но с уменьшением молекулярной активности.Получены данные, свидетельствующие о том, что липиды, создающие микроокружение Na+, К+-АТФазы в биомембранах, характеризуются сравнительно низкими энергиями взаимодействия с ферментом. Существует мнение, что Na+, К+-АТФаза не имеет аннулюса в традиционном понимании этого термина. Вместе с тем исследования с использованием ЭПР-спектроскопии спин-меченных фосфолипидов показали, что фермент оказывает существенное влияние на фосфолипидный состав ближайшего микроокружения в мембране, преимущественно связывая отрицательно заряженные фосфолипиды. Оказалось, что сродство Na+, К+-АТФазы к отрицательно заряженным фосфолипидам в среднем в 4 раза больше, чем к положительно заряженным, и я в 2,5 раза больше, чем к нейтральным. Эта избирательность фермента по отношению к заряду молекул фосфолипидов может быть связана с наличием положительно заряженных боковых цепей аминокислотных остатков во внутримембранном домене Na+, К*-АТФазы, взаимодействующем с полярными головками фосфолипидов из ближайшего микроокружения. Для ряда интегральных мембранных белков обнаружены такие боковые цепи — остатки аргинина и лизина, преимущественно взаимодействующие с отрицательно заряженными фосфолипидами бислоя. Получены доказательства того, что изменение физического состояния липидного окружения Na*, К*-АТФазы под влиянием температуры, содержания в нем холестерина, длины цепи и степени ненасыщенности жирных кислот, входящих в состав фосфолипидов, может влиять на активность этого фермента.
I. J. De Pont et al. (1978) показали, что гидролиз различных фосфолипидов, содержащихся в высокоочищенном препарате Na+, К*-АТФазы из ткани почек крокодила с помощью фосфолипазы С, существенно не влияет на активность фермента. С другой стороны, удаление 60 % фосфолипидов из такого же препарата белка с помощью фосфолипазы А приводило к потере 70 % АТФазной активности, в то время как число уабаин-связываю- щих центров оставалось неизменным. Такой разброс данных о липидной “потребности” Na*, К+-АТФазы объясняется тем, что снижение ферментативной активности обусловлено не только уменьшением содержания фосфолипидов, но и ингибирующим действием жирных кислот, образующихся при гидролизе фосфолипидов фосфолипазами.
Все вышеизложенное свидетельствует о том, что целый рад вопросов, касающихся детального выяснения роли липидного матрикса в функционировании мембраносвязанной Na*', К+-АТФазы, остается недостаточно изученным.
По всей вероятности, Na* , К+-АТФаза находится в тесной пространственной и функциональной взаимосвязи не только с липидами, но и с мембранными белками. Были получены доказательства взаимодействия этого фермента с периферическим белком мембранного скелета — анкирином, установлена его связь с анионным переносчиком — белком полосы 3, выявлено регуляторное влияние на активность АТФазы спектрии-актинового комплекса. Имеются данные, указывающие на возможность образования за счет белок-белковых взаимодействий в мембране эритроцитов сложных мультиферментных комплексов, состоящих из глицеральдегид-3-фосфатдегидрогеиазы, фосфоглицераткиназы, фосфоглицератмутазы и Na+, К'-АТФазы. Между указанными ферментами существует функциональное сопряжение, за счет которого субстраты — 1,3-дифосфоглицерат и АТФ — непосредственно транспортируются от одного компонента системы к другому.
Известно, что Na+, К+-АТФаза может быть функционально связана с такими мембранными системами как, Na+/Ca2'1-обменник, рецептор инсулина, ^-адренорецепторы, сАМР-зависимая протеинкиназа, аденилатциклаза.
Обсуждается вопрос о существовании in vivo пространственной и функциональной взаимосвязи между мембраносвязанными Na4, К+-АТФазой и ацетилхолинэстеразой. Субстрат ацетил- холинэстеразной реакции - ацетилхолин был одним из первых соединений, идентифицированных как нейротрансмиттеры. Он принадлежит к группе нейропередатчиков, вызывающих деполяризацию постсинаптических мембран, которая приводит к возникновению пикового потенциала и торможению активности Na+, К+-АТФазы. Действие ацетилхолина в основном устраняется путем его энзиматического расщепления ацетилхолинэстеразой.
Регуляторные механизмы, в реализации которых, очевидно, участвуют вторичные мессенджеры (например, С а24 и сАМР), гормоны, биогенные амины, нейротрансмиттеры и другие факторы, наименее изучены. Однако не вызывает сомнений, что именно благодаря им происходит более тесное сопряжение натриевого насоса с важнейшими процессами клеточного метаболизма.
Существенное значение для проявления функциональной активности Na+, К+-АТФазы и других векторных ферментов (Са2+-АТФаза, аденилатциклаза, цитохромоксидаза), состоящих из нескольких субъединиц, имеют степень олигомеризации и кооперативные взаимодействия субъединиц или доменов в составе олигомерных молекул, которые находятся под контролем внутренних и внешних факторов. Все вышеперечисленные регуляторные звенья, а именно: взаимодействие физиологических лигандов со специфическими центрами связывания; индуцируемые лигандами взаимодействия доменов внутри субъединиц, отдельных субъединиц, протомеров, олигомерных комплексов; модуляторные эффекты липидного матрикса — лежат в основе краткосрочной (быстрой) или “оперативной” регуляции активности векторных ферментов биомембран при изменении функционального состояния клетки. Этот механизм реализуется за счет электростатических, гидрофобных, ван-дер-ваальсовых взаимодействий, водородных, ковалентных связей, участвующих в поддержании липид-белковых, белок-белковых, липид-липидных взаимодействий (раздел 1.4).
“Долгосрочный” механизм регулирования активности мембранных ферментов реализуется за счет белоксинтезирующей системы клетки и заключается в поддержании оптимального соотношения между скоростью биосинтеза и распада этих ферментов. Он направлен на обеспечение биогенеза необходимого количества функционально активных молекул и олигомерных комплексов векторных ферментов, приходящихся на единицу площади поверхности мембраны. “Долгосрочный” механизм связан с действием различных гормонов, вторичных мессенджеров и других факторов на плазматическую мембрану клетки.
Биосинтез векторных ферментов биомембран на начальных стадиях происходит аналогично синтезу большинства белков эукариотических клеток, однако имеет и ряд особенностей. Так, мембранные белки синтезируются, как правило, в виде неактивных форм (проферментов), которые с помощью селективного протеолиза превращаются в активные формы в ходе посттрансляционной модификации. Вероятно, биосинтез различных субъединиц олигомерных молекул мембранных белков происходит на разных мРНК в шероховатой (гранулярной) эндоплазматической сети. Полагают, что эффективность синтеза мембранных белков значительно повышается за счет объединения отдельных рибосом в полисомы. Однако более детальные исследования, касающиеся выявления важных аспектов биогенеза мембранных белков, остаются не выясненными до настоящего времени.
“Краткосрочный” и “долгосрочный” механизмы регулирования активности мембранных ферментов в реальных условиях in vivo дополняются многочисленными компонентами: функциональным “сопряжением” одного фермента с другими, наличием каскадных механизмов регуляции, “модуляцией” активности белков мембран в результате воздействия физических агентов. В целом процесс регулирования функционирования векторных белков-ферментов биомембран рассматривается как сложное-вза- имодействие “подсистем” универсального регуляторного механизма, обеспечивающее структурно-функциональную интеграцию компонентов мембран и поддержание клеточного гомеостаза (рис. 25). Универсальность основных регуляторных механизмов векторных ферментов биомембран обусловлена сходством их
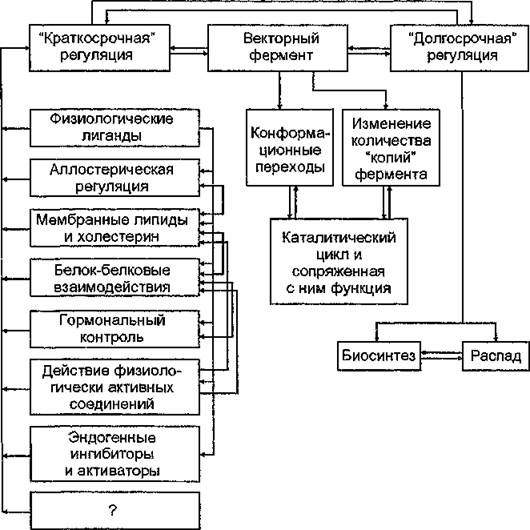
Риє. 25. Схема процессов регуляции активности векторных ферментов биомембран (А. В. Кравцов, И. Р. Алексеенко, 1990)
структурной организации и трансмембранным характером выполняемых ими функций. Вместе с тем остается открытым вопрос о приоритетности того или иного регуляторного механизма и об их взаимодействии в процессе функционирования каждого конкретного фермента. Таким образом, разрабатываемая в настоящее время концепция общности принципов регуляции активности ферментов, выполняющих трансмембранные функции, требует новых экспериментальных доказательств.
2.3.1.