Адсорбционный механизм регуляции метаболизма: понятие о метаболоне, его структура, физиологическое значение образования
В последние годы наряду с “классической” энзимологией, рассматривающей ферментные системы как гомогенный раствор со свободно плавающими ферментами и метаболитами, выделилось направление, основанное на пространственной и структурной организации ферментных систем.
В рамках этого направления разработаны различные методические подходы для решения проблемы надмолекулярной организации метаболических процессов в клетке. Так, целый ряд работ посвящен экспериментальному исследованию. взаимодействия различных структурных компонентов клетки с ферментами важнейших метаболических путей, в частности, гликолиза.Впервые взаимодействие гликолитических ферментов со структурными белками мышц было описано Н. Arnold, D. Pette (1968), которые установили, что на долю F-актина приходится больше связанных ферментов гликолиза, чем на долю миозина, актомиозина и белков стромы. Из ферментов наибольшим сродством к F-актину обладает альдолаза, несколько меньшим — глицеральдегид-3-фос- фатдегидрогеназа. Определенную способность к взаимодействию проявляют также фрсфофруктокиназа, 3-фосфоглицераткиназа, пи- руваткиназа и лактатдегидрогеназа. F. М. Clarke, С. J. Masters (1975) изучили способность ферментов гликолиза образовывать ферментативно-активные комплексы с F-актином и F-актин-—тропомио- зин—тропонином из мышц быка. Они показали, что такие ферменты, как фосфофруктокиназа, альдолаза, пируваткиназа, лактатдегидрогеназа, глюкозо-6-фосфатизомераза, легко ассоциируют с F-актином и нативным тонким: филаментом. Причем наибольшее
сродство эти ферменты проявляют к комплексу F-актин______ тропоми-
озин*—тропонин. 1 риозофосфатизомераза, фосфоглицераткиназа, фосфоглицератмутаза, енолаза и гексокиназа связываются гораздо медленнее. Наилучшие условия ассоциации наблюдались при физиологической концентрации солей - 5,0 ммоль/л КС1 и высокой концентрации миогена — 35 мг/мл.
Это взаимодействие чувствительно к изменению ионной силы: КС1 в концентрации 150 ммоль/л предотвращает образование комплексов.В опытах по экстракции ферментов из грудной мышцы курицы также была показана специфическая адсорбция гликолити- ческих ферментов, особенно глицеральдегид-3-фосфатдегидрогена- зы и лактатдегидрогеназы (J. В. Ehmann, Н. О. Hultin, 1973; R. L. Melrdck, Н. О. Hultin, 1973). Кинетические параметры обоих ферментов при связывании изменялись: максимальная скорость и константа Михаэлиса выше для свободной глицеральдегид-3- фосфатдегидрогеназы; при переходе лактатдегидрогеназы в связанную форму предотвращается ее ингибирование пируватом.
Предположение о существовании на мембране эритроцитов структурно упорядоченного комплекса гликолитических ферментов было высказано еще в 1965 г. D. Е. Green et al. Позднее с использованием метода седиментации на препарате миогена было обнаружено увеличение кажущихся молекулярных масс альдолази, лактатдегидрогеназы, пируваткяназы и фосфофруктокиназы (F. М. Clarke, С. J. Masters, 1973), Полученные результаты подтвердили наличие полиферментных комплексов гликолитических компонентов при физиологических условиях pH и ионной силы.
На основе анализа данных по внутриклеточному распределению ферментов гликолиза в мышечных клетках D. Pette (1975) пришел к заключению о существовании упорядоченного мультиферментного комплекса гликолитических ферментов, адсорбированных на структурных белках мышц. Гистохимическими исследованиями подобный комплекс ферментов был выявлен в изотропной зоне миофибрилл мышечных волокон (Р. Sigel, D. Pette, 1969).
J. Mowbray, V. Moses (1976) обнаружили мультиферментный комплекс, обладающий активностями всех ферментов гликолиза, в растворимой фракции разрушенных механическим путем сферопластов Escherichia coli. F. R. Opperdoes и P. Borst (1977) выявили, что полный набор ферментов гликолиза африканской трипаносомы Trypanosoma brucei упакован в примембранной органелле, названной ими “гликосомой”.
При изучении обратимого связывания ферментов гликолиза в скелетных мышцах было высказано предположение о том, что ассоциация — диссоциация этих ферментов с сократительным аппаратом является регулятором гликоли- тического пути (Т. Р. Walsh et al., 1981).
В ряде работ обсуждается возможный характер пространственной организации комплекса ферментов гликолиза. П. Фридрих предложил гипотетическую схему расположения ферментов в мембране эритроцитов (рис. 20). Идея схемы состоит в наложении последовательно расположенных ферментов центральной части гликолиза на пучок “хвостов” белка полосы 3, имеющих весьма специфичные участки для связывания каждого фермента. Схема учитывает также возможное участие белков цитоскелета (актина) в процессе связывания отдельных ферментов.
Б. И. Курганов и соавт. (1986, 1988) выдвинули гипотетическую модель структур комплекса гликолитических ферментов в скелетных мышцах, а также на внутренней поверхности мембраны эритроцитов. Возможность объединения гликолитических ферментов в комплекс предопределена тем, что их молекулы имеют центры узнавания своих “соседей”, а однозначность сборки достигается тем, что в ней принимает участие якорный белок подложки, на
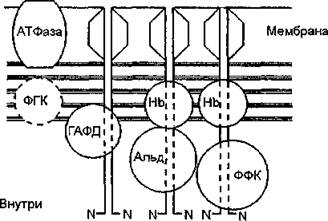
Рис. 20. Гипотетическая схема расположения ферментов в мембране эритроцитов (П. Фридрих, 1986). Димеры В-З-Р (белка полосы 3) — поіру- женньте в мембрану гексагональные структуры. Их N-концы формируют в липидном бислое каналы для анионов и выступают в цитоплазму. ФГК — 3-фосфоглицераткиназа; НЬ — гемоглобин; ГАФД - глицеральдегид-3- фосфатдегидрогеназа; Альд. — альдолаза; ФФК — фосфофруктокиназа
которой формируется комплекс. Комплексы отдельных гликолитических ферментов непрочны, что затрудняет их изучение в растворе. Кооперативность процесса ассоциации приводит к тому, что ассоциация должна протекать по принципу “все или ничего”. Комплекс гликолитических ферментов формируется на тонких нитях изотропной зоны миофибрилл мышечных волокон, образованных F-актином и регуляторными белками — тропомиозином и тропонином, или на димерах белка полосы 3, погруженных в мембрану эритроцитов.
Физиологический смысл формирования комплекса ферментов, участвующих в общем метаболическом пути, на подложке биологической природы состоит в том, что клетка получает возможность регулировать метаболическую систему как единое целое с помощью химических сигналов, воздействующих на “якорный” белок подложки (Б. И. Курганов, 1986).Предполагают, что в скелетных мышцах гликолитическая система стимулируется ионами Са2+ , связывающимися с “якорным” белком подложки — тропонином С. Первый этап сборки комплекса — это, по всей вероятности, посадка фосфофруктоки- назы на “якорный” белок. Остов “ядра” комплекса, так называемое “ядрышко”, образуют в мышечной ткани, кроме фосфофрук- токиназы, альдолаза и глицеральдегидфосфатдегидрогеназа. В состав “ядра” входят глюкозофосфатизомераза, пируваткиназа, лактатдегидрогеназа и фосфоглицераткиназа. Остальные компоненты комплекса (фосфоглицеромутаза, енолаза, триозофосфати- зомераза, глицерол-3-фосфатдегидрогеназа) занимают, очевидно, положения на периферии комплекса (рис. 21).
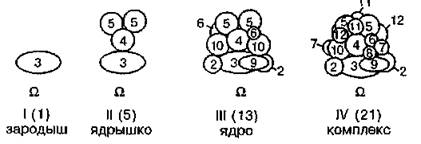
Рис. 21. Этапы сборки комплекса ферментов гликолиза: О — якорный белок подложки; 2 — глюкозофосфатизомераза; 3 — фосфофрук- токиназа; 4 — альдолаза; 5 — глицеральдегидфосфатдегидрогеназа; 6 — фосфоглицераткиназа; 7 — фосфоглицеромутаза; 8 — енолаза, 9 — пи- руваткиназа, 10 — лактатдегидрогеназа; 11 — триозофосфатизомераза; 12 — глицерол-3-фосфатдегидрогеназа. В скобках указано число молекул ферментов в состояниях I—IV комплекса
Сборка гликолитического метаболона на внутренней поверхности мембраны эритроцитов происходит на гексамерах (тримерах димеров) белка полосы 3, имеющих ось симметрии третьего порядка, перпендикулярно к плоскости мембраны. Первым этапом сборки комплекса гликолитических ферментов является адсорбция б-фосфофруктокиназы (ей принадлежит ключевая роль) на олигомерах белка полосы З (Б. И. Курганов, А. Е. Любарев, 1988).
Метаболой содержит тройной набор ферментов, его молекулярная масса составляет 4,510е Да (рис. 22). Метаболой является мобильной структурой и находится в подвижном равновесии со свободными ферментами, которое контролируется клеточными метаболитами (Б. И. Курганов и соавт., 1986). Микроком-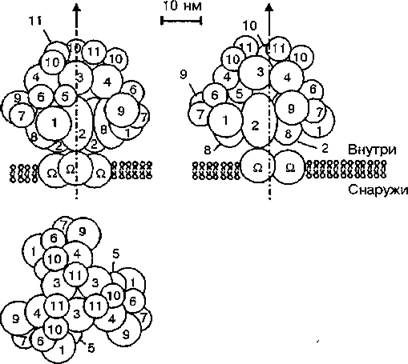
Рис. 22. Гипотетическая структура комплекса ферментов гликолиза, формирующегося на внутренней поверхности мембраны эритроцитов (три проекции) (Б. И. Курганов, А. Е. Любарев, 1988): Q — димер белка полосы 3; 1 — глюкозофосфатизомераза; 2 — 6-фосфофруктоки- наза; 3 — фруктозобисфосфатальдолаза; 4 — глицеральдегидфосфатде- гидрогеназа; 5 — фосфоглицераткиназа; 6 — мультифункциональный фермент, обладающий фосфоглицеромутазной, бис-фосфоглицеромутаз- ной и бис-фосфоглицератфосфатазной активностями; 7 — енолаза; 8 — пируваткиназа; 9 — лактатдегидрогеназа; 10 — триозофосфатизомера- за; 11 — глицерол-3-фосфатдегидрогеназа
партмент имеет вход(ы) для поступления исходных субстратов и выход(ы) конечных продуктов метаболического пути. “Места” выхода АТР — продукта гликолитического пути - в комплексе ферментов гликолиза находятся вблизи фосфоглицераткиназы и пируваткиназы. По-видимому, АТР, продуцируемый фосфогли- цераткиназой, утилизируется Na+, К+-АТФазой.
Таким образом, комплекс гликолитических ферментов, адсорбированный на димерах белка полосы 3, может находиться в контакте с Na+, К+-АТФазной системой и обеспечивать потребности последней в АТР. Взаимодействия ферментов, входящих в состав мультиферментного комплекса, обеспечивают возможность регулирования функциональной активности комплекса как единого целого. Роль факторов контроля отводится в данном случае не интермедиатам метаболического процесса, а внешним факторам - вторичным посредникам, с помощью которых осуществляется оптимальное функционирование мультиферментного комплекса в рамках системы более высокого уровня сложности, т.е.
в клетке. Рассмотрим это на следующем примере. Димер белка полосы 3 эритроцитарных мембран способен взаимодействовать с ионами кальция. Концентрация ионов кальция в эритроцитах составляет 10—20 мкмоль/л. Внутриклеточная концентрация кальция может повышаться в ответ на связывание гормонов и нейромедиаторов соответствующими рецепторами, встроенными в мембрану эритроцитов (см. раздел 2.1.). Изменение степени насыщения якорного белка ионами кальция будет приводить к конформационным изменениям якорного белка и, следовательно, молекул ферментов комплекса, находящихся в непосредственном контакте с якорным белком. В результате изменяются каталитические свойства всех ферментов гликолиза. Кроме того, происходит снятие стеричес- ких затруднений для входа субстратов в микрокомпартмент и дальнейшей химической трансформации в микрокомпартмен- те. Якорный белок — центр управления гликолитическим комплексом — может фосфорилироваться с участием протеинки- наз, которые активируются сАМР — вторичным мессенджером.Следовательно, сборка ферментов, участвующих в общем метаболическом пути, приводит к ослаблению изостерических и аллостерических механизмов и обеспечивает возможность реализации иерархически более “высокого” регуляторного механизма, осуществляющего контроль функционирования метаболической системы как целого. Этот механизм контроля реализуется с участием вторичных мессенджеров, т.е. является механизмом слежения, переключающим функционирование метаболической системы в новый режим в соответствии с сигналами, поступающими от более высоких уровней контроля метаболизма. Механизм контроля функционирования мультиферментного комплекса как целого осуществляется с более высокой скоростью в сравнении с изостерически- ми и аллостерическими механизмами регуляции.
Мультиферментные комплексы, формирующиеся на подложках биологической природы, являются мобильными образованиями, которые находятся в равновесии со свободными ферментами. Это обстоятельство определяет возможность эффективного осуществления и взаимодействия в клетке и механизмов гомеостаза, и механизмов слежения.
Ферменты, связанные общими метаболитами или коферментами, находятся в метаболоне рядом друг с другом. Это обеспечивает эффективное продвижение интермедиатов по конвейеру активных центров в микрокомпартменте, образующемся при сборке метаболона (Б. И. Курганов, А. Е. Любарев, 1988).
На основе данных о пространственной организации метаболических процессов в цитоплазме эукариотической клетки А. Г. Рязанов и А. С. Спирин (1989) построили эстафетную модель работы ферментов в клетке, которая объясняет молекулярный механизм компартментализации. Модель основана на трех постулатах: ассоциации ферментов с субклеточными структурами, передачи метаболитов между ферментами без диффузии в растворе и десорбции ферментов с поверхностей внутриклеточных структур в процессе каждого акта катализа (рис. 23). Важным преимуществом механизма “эстафеты у поверхности” является то, что он приводит к
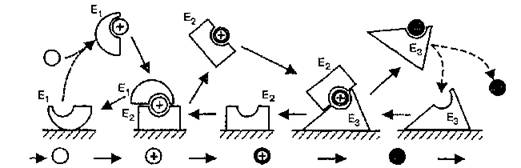
Рис. 23. Схема эстафетной передачи метаболитов между ферментами у поверхности внутриклеточной структуры
концентрированию и компартментализации ферментов данного метаболического пути вблизи места потребления конечного продукта. Те или иные модификации ферментов или поверхности, изменяющие их взаимное сродство, создают возможности нового механизма регуляции метаболических процессов за счет пространственного перераспределения ферментов в клетке.
Концепция метаболической регуляции посредством динамической сборки и разборки ферментных комплексов обсуждается также в работе J. Ovadi (1988).
J. Batke (1989) предложена схема процессов взаимодействия между ферментами гликолиза в мышцах (рис. 24).
На основании анализа результатов исследования взаимодействия ферментов цикла трикарбоновых кислот (ЦТК), связанного со множеством анаболических и катаболических процессов, с внутренней мембраной митохондрий, а также белок-белковых взаимодействий А. Е. Любаревым и Б. И. Кургановым (1987) разработана модель, описывающая пространственную структуру комплекса ферментов этого центрального метаболического пути. Предположение о существовании комплекса ферментов ЦТК ими было высказано с учетом следующих экспериментальных дан-
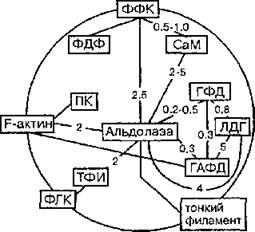
Рис. 24. Схема взаимодействия между ферментами гликолиза в мышцах: ФФК — фосфофруктокиназа; ФДФ — фруктозо-1,6-дифосфатаза; ГАФД — глицеральдегидфосфатдегидрогеназа; ФГК — фосфоглицераткиназа; ПК — пируваткиназа; ЛДГ — лактатдегидрогеназа; ТФИ — три- озофосфатизомераза; ГФД — глицерол-3-фосфатдегидрогеназа; СаМ —- кальмодулин. Линиями обозначена возможность комплексирования in vitro, цифрами представлены значения Kd в микромолях
ных и теоретических расчетов: концентрация белка в матриксе митохондрий, где локализованы ферменты ЦТК, составляет 40 вес. %; в этих условиях затруднена диффузия молекул ферментов и их кофакторов; среднее расстояние между внутренними поверхностями внутренней мембраны митохондрий сердца составляет 15—30 нм.
Авторы считают, что комплекс ферментов ЦТК представляет собой гексамер из шести идентичных асимметричных субъединиц, каждая из которых содержит тетрамер транссукцинилазы, по одной молекуле остальных ферментов и “якорный” белок. Комплекс “зажат” между противоположными поверхностями внутренней мембраны митохондрий, причем с мембранами контактируют все ферменты, за исключением а-кетоглутаратдегидроге- назы и аспартатаминотрансферазы. В качестве “якорных” белков выступают интегральные белки внутренней митохондриальной мембраны, в том числе сукцинатдегидрогеназа. Они обеспечивают сборку комплекса ферментов ЦТК на мембране, высота которого вдоль оси симметрии третьего порядка составляет 20 нм, а диаметр — 50 нм. Молекулярная масса (без учета сукцинатде- гидрогеназы) равна 8 мДа. Структура предложенной модели допускает возможность взаимодействия этого комплекса с другими ферментами и мультиферментными ансамблями, с которыми он связан общими метаболитами, и, прежде всего, с комплексами цепи переноса электронов внутренней мембраны митохондрий. Кроме того, метаболой ЦТК должен взаимодействовать с другими ферментами матрикса. Для митохондрий сердца в качестве таких белков выступают пируватдегидрогеназный комплекс и ферменты (3-окисления жирных кислот, поставляющие в ЦТК аце- тилкофермент А.
Комплекс ферментов ЦТК — мобильная структура, находящаяся в равновесии со свободными ферментами матрикса. Обратимое равновесие между свободными и связанными ферментами контролируется целым рядом факторов: концентрациями определенных метаболитов в матриксе митохондрий (цитрата, окса- лоацетата, кофермента А, ацетилкофермента A, Mg2+-ATP и др.), концентрацией белка, энергетическим состоянием митохондрий. Взаимодействия ферментов, входящих в состав метабол она ЦТК, обеспечивают функционирование этого надмолекулярного комплекса ферментов как целостной, кооперативно действующей системы.
В то же время нет убедительных доказательств существования надмолекулярного комплекса какого-либо метаболического пути или его фрагмента. Прямым подтверждением существования метаболона (надмолекулярного комплекса ферментов, катализирующих последовательные стадии ферментной системы) может быть либо его выделение, либо реконструкция in vitro и экспериментальное доказательство туннелирования между ферментами-соседями (Г. Л. Ермаков, 1993). Такие попытки предпринимаются, например, для ферментов гликолиза, цикла Кребса и цикла мочевины.
Таким образом, в настоящее время широко обсуждается проблема пространственно-структурной организации ферментных систем. Вместе с тем отсутствуют исчерпывающие данные, касающиеся изучения фермент-ферментных, фермент-мембранных взаимодействий, взаимосвязи физико-химических характеристик белков с их способностью образовывать надмолекулярные комплексы, выяснения роли цитоскелета клетки в организации клеточного метаболизма. Кроме того, схемы надмолекулярной организации компонентов метаболических систем требуют экспериментальных доказательств.
2.3.3.