ИССЛЕДОВАНИЕ РОЛИ АКТИВНЫХ ФОРМ КИСЛОРОДА В ПРОЦЕССАХ УФ-МОДИФИКАЦИИ ЛАКТАТДЕГИДРОГЕНАЗЫ[4]
В настоящее время возрос интерес исследователей к проблеме окислительного стресса. Ключевую роль в развитии окислительного повреждения играют активные формы кислорода (АФК): синглетный молекулярный кислород Ю.,, супероксидный анион- радикал, пероксид водорода Н2О2, гидроксильный радикал ОН’, пергидроксильный радикал, гипогалоиды (см.
разделы 3.2, 3.3). Образование АФК и оксидативная модификация макромолекул — нормальные и важные регуляторные процессы, участвующие в синтезе эйкозаноидных гормонов (простаноидов и лейкотриенов) и иодтиронинов, в воспалительных и иммунологических реакциях. В норме организм имеет мощную антиоксидантную (защитную) систему, но при различных патологических состояниях и действии экстремальных факторов (избыток О2, активация нейтрофилов и фагоцитов, избыток гема, ионов Ре2+ и Сн2+, ионизирующее и УФ-излучение, некоторые ксенобиотики) необходимо предусматривать дополнительные меры, повышающие устойчивость макромолекул (белков и нуклеиновых кислот) к оксидативной модификации и, в частности, фотоокислению.Существенную роль в процессах темнового и фотоокисления биологических структур играют синглетный кислород и радикал гидроксила, обладающие наибольшим модифицирующим эффектом. Одним из широко используемых методов исследования роли тех или иных активных интермедиатов в процессе УФ-мо- дификации белка является элиминирование их при помощи специфических акцепторов и тушителей.
Рассмотрим более подробно результаты исследования фотоиндуцированных изменений структурно-функциональных свойств изоферментов лактатдегидрогеназы из различных источников в интактном состоянии и в присутствии ряда химических соединений, способных взаимодействовать с активными формами кислорода.
Каталитическая активность растворов изофермента М4 ЛДГ (0,2-Ю’8 моль/л) из скелетных мыщц свиньи при действии УФ- света (240—390 нм) в дозах 0,15—4,53 кДж/м2 снижается с ростом дозы облучения (В.
Г. Артюхов и соавт., 1997, В. Г. Артю-хов, М. А. Наквасина, 1991). NADH (2,4-10 ~7 моль/л) оказывает фотосенсибилизирующее действие по отношению к молекулам фермента при УФ-облучении их смеси. Падение каталитической активности ЛДГ происходит под влиянием УФ-излучения, поглощаемого как апоферментом (А. < 280 нм), так и коферментом (Х> 260 нм). По-видимому, в процесс фотоинактивации ЛДГ вносят вклад преимущественно фотохимические превращения апо- белка, связанные с поглощением энергии УФ-света остатками ароматических (триптофана, тирозина — входит в активный центр ЛДГ, фенилаланина) и серосодержащих аминокислот белка. Присутствие указанных аминокислотных остатков в молекуле мышечной изоформы фермента было продемонстрировано при исследовании ее аминокислотного состава (табл. 14).
Содержание отдельных аминокислот в молекуле лактатдегидрогеназы
Таблица 14
| Аминокислоты | Количество, % на 100 г белка | Аминокислоты | Количество, % на 100 г белка |
| Asp | 7,820 | Met | 3,693 |
| Thr | 4,250 | lie | 4,819 |
| Ser | 3,294 | Leu | 10,579 |
| Glu | 9,640 | Туг | 3,793 |
| Pro | 6,273 | Phe | 4,078 |
| Gly | 4,006 | His | 2,937 |
| Ala | 4,092 | Trp | 2,068 |
| "Полуцистин" | 1,141 | Lys | 9,610 |
| Val | 8,626 | Arg | 6,388 |
Наблюдаемые при воздействии УФ-света нарушения структурного состояния и функциональной ■активности ЛДГ являются результатом разворачивания белковой глобулы, что находит отражение в изменении оптических и гель-хроматографических свойств УФ-облученного фермента.
Продуктом фотодеградации остатков триптофана являются кинуренины и формилкинуренины. Показано, что производные триптофана способны выполнять функцию фотодинамических сенсибилизаторов, в частности, генерировать синглетный кислород. В качестве источника *О2 рассматривают процессы тушения кислородом триплетных состояний ароматических или карбонильных соединений, которые входят в состав макромолекул или присутствуют в них в виде примесей. На потенциальную возможность генерации ’О, в реакции триплетных возбужденных состояний белков указывает И. И. Сапежинский (1979). Образование синглетного кислорода может происходить в короткоживущих комплексах с переносом заряда: ароматический углеводород (аминокислотный остаток) — кислород.
С целью получения дополнительной информации о механизме фотопревращений ЛДГ необходимо исследовать УФ-индуци- рованные изменения структурно-функциональных свойств и кинетики фотоинактивации различных изофермептов ЛДГ в широком спектральном диапазоне УФ-излучения для выяснения возможного участия модифицированных триптофаиилов в качестве «внутримолекулярных аутосеисибилизаторов». Для изучения роли активных кислородных метаболитов в процессах УФ- превращений белка используют их специфические акцепторы и тушители: азид натрия, D-маннит, (3-каротин, гистидин, серотонин. Применяемые концентрации вышеназванных соединений не должны оказывать заметного влияния на уровень активности нативного фермента.
Распространенным тушителем синглетного кислорода является азид натрия, защитное действие которого было продемонстрировано по отношению к сенсибилизированному окислению различных органических субстратов. Константа скорости его реакции с ‘Оа — 2-Ю8 л-(моль-с)"1, 2-10® л-(моль-с)"1. Азид натрия тушит триплетные состояния возбужденных молекул, но константа скорости данной реакции в последнем случае на 2 порядка ниже. Для подбора концентрации NaN3 можно использовать предположение о простом конкурентном перехвате синглетного кислорода в исследуемой модельной системе и в соответствии с известными'константами скорости взаимодействия 'О, с компонентами этой системы.
Для ЛДГ такая константа не определена, но в первом приближении можно принять ее значения равными 10®— 10го л-(моль-с)"1 (И. И. Сапежинский, 1988). Суммируя известные литературные данные (Б. Рэнби, Я. Рабек, 1978, В. Я. Шляпин- тох, 1979, X. Фут, 1979, Н. А. Шинкаренко, В. Б. Алесковский, 1981, Ю. А. Владимиров, А. Я. Потапенко, 1989), взаимодействие синглетного кислорода с возможными субстратами реакции может быть представлено с помощью следующей упрощенной формально-кинетической схемы: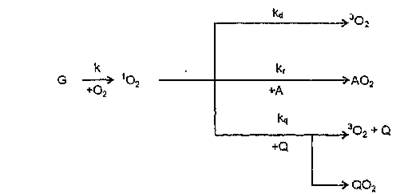
где G —' источник, генерирующий *О2; kd — константа дезактивации ‘О2 растворителем; kr — константа скорости реакции 1О2 с акцептором (окисляемым веществом); к — константа скорости тушения (или химического взаимодействия) с потенциальным протектором,
В присутствии двух конкурирующих за *О2 соединений (А и Q) квантовый выход окисления акцептора определяется по уравнению

Величина второго слагаемого в правой части неравенства будет пренебрижимо мала по сравнению с первым до значений концентраций фермента -10“° моль/л. Следовательно, в этом случае
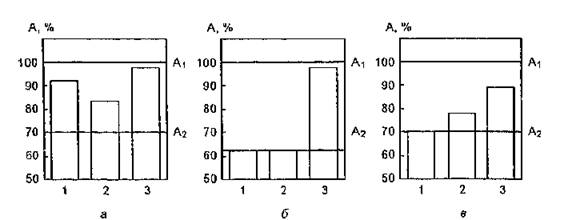
Рис. 54. Изменение фоточувствительности ЛДГ в присутствии азида натрия (я), p-каротина и D-маннита (5), гистидина (я). По оси абсцисс — концентрации модификаторов, моль/л; по оси ординат — ферментативная активность, % . — активность нативного фермента, А2 — УФ-облу-
ченного фермента. Концентрации модификаторов: а) 1 — 6,6-10'“ моль/л; 2 —• 6,6-Ю"4 моль/л; 3 — 3,2-10'3 моль/л; б) 1 — 3,3-10'“ моль/л (Р-каротин); 2 — 5,5-Ю’5 моль/л (D-маннит); 3 — Р-каротин + D-малнит; в) 1 — 1,3-10"3 моль/л; 2 — 1,3-10'2 моль/л; 3 — 3.3-10'2 моль/л
концентрация азида натрия для проявления фотопротекторного эффекта должна превышать 1,6-1О'3(~4) моль/л.
Эти теоретические расчеты были подтверждены результатами экспериментов по изучению УФ-чувствительности (доза облучения — 2,27 кДж/м2) изофермента М4 ЛДГ (2-Ю'8 моль/л) в присутствии азида натрия в концентрациях 6,6-Ю'5; 6,6-Ю'4; 3,2-Ю"3 моль/л (рис. 54,я). Из анализа рисунка следует, что NaN3 в концентрации 3,2-10"3 моль/л практически полностью восстанавливает ферментативную активность исследуемого белка. При изучении спектрально-люминесцентных характеристик ЛДГ (1О'° моль/л) после ее облучения в свободном состоянии и в присутствии азида натрия (3,2-10~3 моль/л) выявлено, что воздействие УФ-света на смесь фермент-азид натрия приводит к восстановлению до уровня контрольного образца величин интенсивности люминесценции модифицированного белка в его максимуме (340 нм) и светопоглощения в минимуме (250 нм) и в более длинноволновой области (>290 нм), что связано, вероятно, с защитой хромофорных групп ЛДГ от действия УФ-излучения.
На рис. 55, а показаны кривые зависимости остаточной ферментативной активности мышечной изоформы ЛДГ от времени облучения. Для количественного описания данного процес-
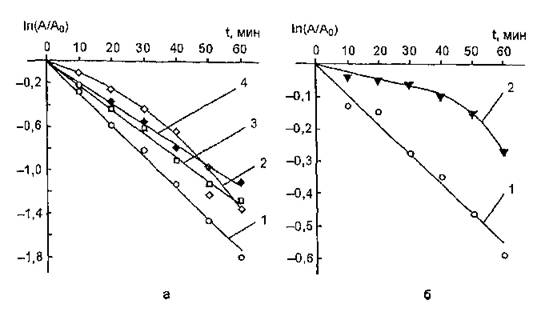
Рис. 55. Кинетические кривые фотоинактивации лактатдегидрогеназы из мышц (а) и сердца {5} свиньи в присутствии некоторых фото- протекторов: а) 1 — нативная ЛДГ; ЛДГ, УФ-облученняя в присутствии модификаторов: 2 — азида натрия (3,2-lQ"s моль/л); 3 — D-маннита (5,5-10‘s моль/л); 4 — серотонина (1-Ю'5 моль/л); б) 1 — нативная ЛДГ; 2 — ЛДГ, УФ-облучеиная в присутствии азида натрия (3,2-10*3 моль/л)
са мы использовали константы фотоинактивации (kf), рассчитанные по тангенсу угла наклона линейной анаморфозы исходной кривой «доза — эффект» в координатах [ln(A/A0); t]. Для нативной ЛДГ kf оказалась равной 5,39-Ю"4 с-1.
Экспоненциальный характер кривой фотоинактивации свидетельствует об отсутствии процессов реактивации УФ-модифицированного белка и позволяет отнести последовательность фотохимических реакций ЛДГ к категории мономолекулярных реакций первого порядка. Время полупревращения ЛДГ (период, в течение которого каталитическая активность белка уменьшается на 50 %) в этом случае составит: t1/2= In 2/k = 0,693/k = 1286 с = 21 мин. При концентрации фермента 0,2-Ю'8 моль/л («преддиссоциационной») форма кинетической кривой фотоинактивации не изменяется, однако к, существенно увеличивается и становится равной 3,05-10"3 с"1, а t = 227,2 с = 3,8 мин. Следовательно, УФ-чувствительность ЛДГ возрастает при разведении ее растворов.Добавление азида натрия (3,2-10‘3 моль/л) при облучении раствора изоформы М4 ЛДГ (2-Ю-8 моль/л) вызывает частичное восстановление каталитической активности фермента. Однако
дозовая зависимость описывается прямой, а в полулогарифмических координатах — квадратичной функцией (см. рис.55, а). Это свидетельствует о постоянстве скорости инактивации ЛДГ в изучаемом диапазоне. Таким образом, протекторный эффект NaN3 на разных стадиях фотомодификации фермента неодинаков. Защитное действие азида натрия может быть обусловлено тушением ‘Og, а также его комплексированием с белковой макромолекулой и, возможно, образованием внутримолекулярных сшивок некоторых функциональных групп ЛДГ, что индуцирует повышение фоторезистентности зафиксированной конформации биополимера. Не исключено также тушение азидом натрия возбужденных состояний белка и его взаимодействие со свободными радикалами, образующимися в процессе УФ-модификации молекул фермента.
На рис. 55, б представлена зависимость «доза — эффект» для изоформы ЛДГ из сердца свиньи в полулогарифмических координатах. Эта зависимость удовлетворительно описывается прямой. Константа скорости инактивации фермента составляет 1,41-lG"4 с"1 и t1/2=1690 с = 28 мин, что позволяет сделать вывод о меньшей фоточувствительности изофермента Н4 по сравнению с М4. При добавлении к раствору ЛДГ до облучения азида натрия остаточная ферментативная активность оказалась выше таковой для облученной в соответствующих дозах свободной изоформы (см. рис. 55, б).
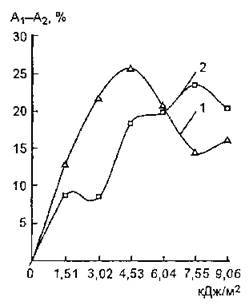
Рис. 56. Сравнение протекторного эффекта азида натрия по отношению к сердечной и мышечной изоформам ЛДГ при различных дозах УФ-облучения: 1 — протекторный эффект NaN3 для мышечной изоформы; 2 — для сердечной изоформы. По оси ординат отложена разность между уровнями ферментативной активности ЛДГ, УФ-облученной в смеси с азидом натрия (Ах) и в свободной форме (А2)
На рис, 56 представлена зависимость протекторного эффекта азида натрия от дозы УФ-облучения по отношению к мышечной и сердечной изоформам ЛДГ. Анализ полученных данных позволяет сделать заключение о неоднонаправленном характере действия модификатора: максимальное защитное действие азида натрия наблюдается для ЛДГ (М() при дозах облучения 3,02—6,04 кДж/м2, а по отношению к ЛДГ (Н4) — 6,04—9,06 кДж/м2. Если предположить, что основной вклад в фотопротекторный эффект NaN3 вносит процесс тушения ‘О,,, то этот активный интермедиат оказывает влияние на изменение ферментативной активности изоформ ЛДГ при вышеуказанных дозах облучения.
Наиболее широко распространено представление о том, что дезактивация ’О2 p-каротином, взаимодействующим с ним преимущественно по физическому механизму, связана с переносом энергии от синглетного кислорода на низколежащий триплетный уровень каротина. Обсуждается вопрос о вкладе в рассматриваемый процесс комплекса с переносом заряда между 1О2 и каротином. Константа скорости дезактивации синглетного кислорода близка к диффузионной и составляет для СС1( 7-Ю9 л-(моль-с)'1. Гистидин относят к типу акцепторов его дезактивация осуществляется химическим путем. Константа скорости к для этой аминокислоты составляет 5-Ю7 л-(моль-с)-1 в метаноле; в D2O — 108 л-(моль-с)”1. В реакцию с *О2 вступает только непротонированная форма гистидина. Из анализа рис. 54, б следует, что Р-каротин в концентрации 3,3-10~6 моль/л оказывает защитное действие по отношению к молекулам ЛДГ (М4): активность УФ-облученного в дозе 2,27 кДж/м2 белка восстанавливается на 18 % по сравнению с таковой в отсутствие протектора. Гистидин в концентраций З.ЗПО'2 моль/л снижает степень инактивации ЛДГ на 19 % (см. рис, 54, в), что согласуется с данными, полученными при использовании р-каротина.
Облучение ЛДГ в присутствии D-маннита, являющегося акцептором ОН-радикалов, сопровождается снижением ее фоточувствительности: константа инактивации составила 4,0-10 '1 с 1, a t1/2 = 1732 с = 29 мин (рис. 55, а), Дозовая кривая имеет экспоненциальный характер. При изучении сочетанного воздействия Р-каротина и D-маннита на уровень активности УФ-облученной ЛДГ выявлен четко выраженный эффект аддитивности действия этих агентов (рис. 54, в). Полученные результаты свидетельствуют о том, что их протекторное действие связано с реализацией различных каналов защиты белка от УФ-повреждения: элиминированием Ю2 и ОН-радикалов.
При УФ-облучении ЛДГ (МД из скелетных мышц свиньи в присутствии серотонина установлено его фотопротекторное действие по отношению к молекулам этой изоформы: kf =3,5-10“4 с-1 и t = 1980 с = 33 мин (см. рис. 55, а).
На рис. 57 показаны фотоиндуцированные изменения функциональных свойств ЛДГ (изоферменты Н4, Н3М, Н2М2) эритроцитов крови человека после УФ-облучения в присутствии серотонина. Введение в облучаемую систему экзогенного агента в концентрации 10~8 моль/л приводит к проявлению его защитного действия только при использовании максимальной дозы УФ-света (4,53 кДж/м2), вызывающей наиболее глубокие изменения в структуре молекулы фермента и приводящей к падению его активности на 35 % по сравнению с контрольным образцом. Увеличение концентрации серотонина до 10~7 моль/л индуцирует резкое усиление его фотопротекторного эффекта, наиболее ярко проявляющегося при облучении ЛДГ в дозах 1,51 и 4,53 кДж/м2. При использовании биогенного амина в концентрации 10~° моль/л отмечается снижение его защитного действия при воздействии УФ-излучения в дозах 1,51 и 4,53 кДж/м2 по отношению к этому показателю при концентрации серотонина 10-7 моль/л. Протекторный эффект 5-окситриптамина может быть связан либо с акцепцией различных активных интермедиатов, в том числе и ^2, либо с образованием комплекса фермент — биогенный амин, более фоторезистентного, чем свободный белок.
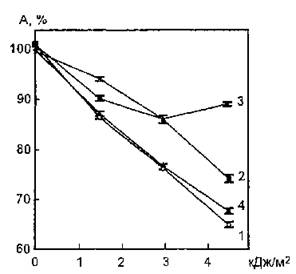
Рис, 57. Зависимость уровня каталитической активности лактатдегидрогеназы эритроцитов человека в свободном состоянии и в присутствии серотонина от дозы УФ-облучения: 1 — УФ-облучение без модификатора; УФ-облучение в присутствии серотонина: 2 — в концентрации 1О’в моль/л, 3— 10"7 моль/л, 4— 10"8 моль/л
С целью детализации представлений, касающихся выяснения механизма защитного действия серотонина по отношению к молекулам ЛДГ, при помощи метода ИК-спектрофотометрии были исследованы особенности вторичной структуры изофермента М4 из скелетных мышц свиньи в нативном состоянии и в присутствии биогенного амина.
Анализ соотношения интенсивностей поглощения в полосах амид I и амид II на частотах, являющихся характеристическими для отдельных элементов вторичной структуры, свидетельствует о том, что присутствие биогенного амина индуцирует увеличение доли беспорядочной структуры в молекуле фермента (табл. 15). При этом отмечаемый прирост ее реализуется за счет как а-спи- ральных участков, так и p-складчатых слоев. В случае соотношения молекул ЛДГ и экзогенного модификатора 1:1 наблюдаемый эффект в большей степени обусловлен участием а-спиралей, входящих в состав белка.
Таблица 15
Соотношение полос поглощения, характеризующих основные типы вторичной структуры белковой молекулы, для лактатдегидрогеназы в свободном состоянии и в присутствии серотонина
| Соотношение интенсивностей полос поглощения | Исследуемые образцы | ||
| Нативный белок | ЛДГ + серотонин в соотношении молекул 1:1 | ЛДГ + серотонин в соотношении молекул 1:10 | |
| 1656 (беспорядочная структура): 1650 (а-спираль) см'1 | 1,423 ±0,092 | 1,406 ± 0,021 | 1,596 ± 0,107 |
| 1535 (беспорядочная структура): 1516 (а-спираль) см'1 | 1,488 ±0,092 | 2,297 ± 0,078* | 1,658 ± 0,109 |
| 1685 (Р-складки): 1656 (беспорядочная структура) см'1 | 0,926 ± 0,007 | 0,932 ± 0,028 | 0,752 ± 0,040* |
| 1550 (Р-складки): 1535 (беспорядочная структура) см"1 | 1,342 ±0,058 | 1,072 ± 0,029* | 1,160 ± 0,033* |
| 1650 (а-спираль): 1685 (р-складки) см'1 | 0,746 ±0,050 | 0,756 ± 0,029 | 0,754 ±0,046 |
| 1516 (а-спираль): 1550 (Р-складки) см'1 | 0,538 ±0,029 | 0,407 ± 0,009* | 0,535 ±0,019 |
* Отличие от контроля статистически достоверно.
Изменение спектральных характеристик фермента удается обнаружить и на других участках спектра. При соотношении молекул ЛДГ и серотонина 1:1 статистически достоверно уменьшается интенсивность поглощения образцов на частотах 1468, 1420 см-1 по сравнению с нативным белком; отмечается появление нового максимума на частоте 1502 см-1.
Повышение концентрации экзогенного модификатора в 10 раз сопровождается снижением интенсивности поглощения на частоте 1432 см'1. Вместе с тем в области 1440—1380 см'1 регистрируется выраженный пик с максимумом при 1404 см'1. Для нативного белка поглощение в этом участке незначительно, а при соотношении компонентов изучаемой системы 1:1 практически отсутствует. ИК-спектр свободного фермента характеризуется наличием широкой полосы поглощения в области 1370—1180 см-1, Добавление к его раствору серотонина приводит к ее сглаживанию и при соотношении молекул ЛДГ — биогенный амин 1:10 в данном диапазоне отмечается практически монотонное убывание светопропускания образца.
Таким образом, выявлен выраженный фотопротекторный эффект серотонина по отношению к молекулам изоферментов Н4, Н3М, Н2М2 ЛДГ эритроцитов человека, обусловленный образованием комплекса ЛДГ — серотонин, формирование которого затрагивает вторичную структуру белка. Возможно, комплексиро- вание биогенного амина с молекулами фермента происходит и в растворе. Однако полученные результаты не исключают и вероятность дезактивации серотонином АФК, в том числе
Динамический комплекс с молекулами ЛДГ способен образовывать и его кофактор NADH, что может существенно изменять фоточувствительность фермента. С целью расширения представлений, касающихся выявления механизма УФ-модификаций ЛДГ в присутствии кофермента, были проведены эксперименты по изучению фотостабильности молекул ЛДГ (М4) из скелетных мышц свиньи в условиях различной занятости ее активных центров этим кофактором. Степень занятости активных центров фермента NADH рассчитывали по формуле (Ч. Кантор, П. Шиммел, 1985):
Y_ [NADH]
Kdis+[NADH]’
где Y — степень насыщения белка лигандом; [NADH] — концентрация кофактора; Kdis — константа диссоциации комплекса белок — лиганд (в качестве КДз использовали значение константы Михаэлиса 2,5-10"° моль/л для NADH по отношению к изоформе М4) (С. Е. Северин, Г. А. Соловьева, 1989) .
Установлено, что предварительное добавление NADH (4-Ю'8 моль/л) к ЛДГ (10~8 моль/л), УФ-облучаемой в дозе 3,02 кДж/м2, не вызывает статистически достоверных изменений степени инактивации свободного белка (32 %). Y в данном случае — 0,16 %. При концентрации NADH 4-Ю"7 моль/л (Y = 1,57 %) обнаруживается эффект сенсибилизации (13 %). Фотомодификация ЛДГ в присутствии кофермента в концентрациях 4-Ю*6 (Y = 13,8 %) и 2,5-10*° моль/л (Y=50 %) приводит к регистрации протекторного эффекта NADH по отношению к ферменту: степень УФ-инактивации белка в этом случае соответственно на 9,3 и 16,0 % ниже уровня активности ЛДГ, облученной без кофактора.
Путем одноэлектронного окисления NADH и при восстановлении NAD+ может образовываться радикал NAD’, взаимодействующий с кислородом с образованием супероксидного анион- радикала О/, поражающего белковую глобулу. Вместе с тем восстановленная форма кофактора обладает способностью дезактивировать 1О2: константа скорости тушения синглетного кислорода в системе CH3CN : D2O (4:1) — 7,5-Ю7 л-(моль-с)*1; в D2O — 2,1-Ю7 л-(моль-с)'1. Следовательно, значение ее для NADH составляет 1,7-10*4 — 1,5-10*2 моль/л. NADH способен взаимодействовать с О/, причем выявлено, что его окисление супероксидом идет быстрее, если кофактор образует.комплекс с ЛДГ: константа скорости окисления NADH в последнем случае по меньшей мере на 4 порядка выше таковой для свободного лиганда. Окисление NADH в активном центре фермента, по-видимому, должно происходить по цепному механизму и существенным образом изменять химические свойства кофактора.
Спектр поглощения комплекса ЛДГ (2-Ю-8 моль/л) — NADH (2-Ю*8 моль/л) характеризуется двумя полосами поглощения с Хгаш[ при 268 и 340 нм. Присутствие полосы с А,зпах 268 нм в спектре поглощения изучаемой системы можно объяснить индукцией структурных изменений молекулы апофермента путем присоединения кофактора. D. Scherr и соавт. (1973) исследовали спектральные свойства двойных комплексов, образующихся при взаимодействии аналогов или фрагментов NADH с ЛДГ из сердца свиньи. Комплексообразование вызывало спектральные изменения в области 280 нм, что указывает на участие в этом процессе ароматических аминокислот. Авторы постулировали гидрофобный характер взаимодействия адениновой части коэнзима с апо- белком.
Таким образом, суммарный регистрируемый эффект изменения каталитической активности ЛДГ при ее УФ-облучении будет зависеть от соотношения процессов сенсибилизации и протекторного эффекта NADH. Защитное действие кофермента (Y = 13,8 и 50 %) может быть обусловлено реализацией следующих возможных процессов:
1) формированием более фоторезистентной конформации ЛДГ в комплексе с NADH по сравнению с апоферментом;
2) миграцией энергии с возбужденных аминокислотных остатков белка (в частности, триптофана) на кофермент;
3) акцептированием АФК (Ю,, и О2Д);
4) экранирующим эффектом NADH в области длин волн 240— 280 нм.
Фотосенсибилизирующий эффект NADH по отношению к ЛДГ, по-видимому, объясняется образованием при УФ-облучении белка сольватированного электрона, акцептируемого NAD+, в результате чего в присутствии О2 возможен циклический процесс перехода NAD+ — NAD' с генерированием О2~ и других активных интермедиатов (NAD-, Н2О2). Кроме того, вследствие конформационных переходов протомеров ЛДГ при фотоокислении NADH могут формироваться субъединицы, занятые одновременно восстановленной и окисленной формой кофактора, что существенно изменяет характер УФ-превращений, индуцируемых в системе фермент — кофермент.
Изучение роли АФК в процессах фотомодификации белка существенно дополняют исследования фотосенсибилизированного метиленовым голубым (МГ) фотоокисления изофермента М4 ЛДГ. Механизм действия красителя связан с генерацией 1О2 или непосредственным взаимодействием возбужденной молекулы МГ с молекулой белка. Преобладание в системе тех или иных процессов зависит от соотношения концентраций биополимера и красителя в облучаемой системе.
Из анализа данных, представленных на рис. 58, а, следует, что облучение красным светом изофермента М4 (10"8 моль/л) в течение 15 мин с МГ (Ю-7 моль/л) не вызывает статистически достоверных изменений функциональных свойств ЛДГ по сравнению с уровнем активности белка, облученного в отсутствие красителя.
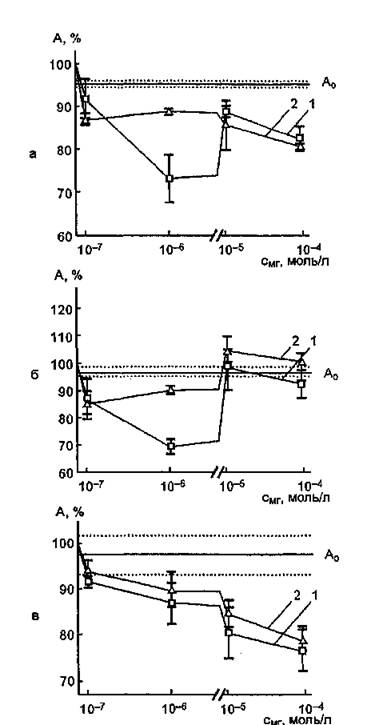
Рис, 58, Зависимость ферментативной активности ЛДГ (М4), облученной красным светом в присутствии азида натрия (3,2-10~3 моль/л), от концентрации метиленового голубого: а — концентрация фермента— 10-8 моль/л; б — 10"° моль/л; в —10“7 моль/л; Ао — активность фермента, облученного в течение 15 мин красным светом без модификаторов; 1 — ЛДГ; 2 — ЛДГ+NaNg
Добавление МГ в концентрациях 10Чі, 10^ и 10"* моль/л сопровождалось эффектом сенсибилизации: активность ЛДГ снижалась на 26,7±6,0; 11,0±1,8 и 17,4±2,6% соответственно. В присутствии NaN8 остаточная активность фермента после его облучения с МГ (10_в моль/л) составляет 87,8±3,6%, что свидетельствует об участии *О2 в процессе фотоинактивации ЛДГ (10-8 моль/л).
При использовании растворов фермента (10_в моль/л) максимальный сенсибилизирующий эффект МГ обнаруживается при его концентрации КР моль/л и составляет 30,0±4,0 %, а остаточная активность белка, облученного в присутствии азида натрия, равна 90,3±2,4% (рис. 58, б). Аналогичные результаты были получены при исследовании фотосенсибилизированного окисления сердечной изоформы ЛДГ с участием МГ моль/л).
Для ЛДГ (1О'Т моль/л) в присутствии МГ характер изменения функциональных свойств ЛДГ от концентрации красителя при облучении их смеси оказался иным: активность фермента убывает с ростом концентрации МГ, а степень инактивации достигает максимальной величины (23,2±4,5%) при концентрации сенсибилизатора 10"* моль/л (рис. 58, в). При добавлении в систему азида натрия значения исследуемого параметра не отличались от таковых для ЛДГ, фотомодифицированной в свободном состоянии.
Взаимодействие фермента с синглетным кислородом может осуществляться при относительно высоких концентрациях белка, так как 1О2 характеризуется малым временем жизни. Этим, вероятно, объясняется отсутствие протекторного действия NaNa для низких концентраций фермента. При определенных концентрациях МГ (10*в и 10~* моль/л) возможно тушение его триплетного состояния своими же молекулами или взаимодействие их с *0,,; этот поражающий агент не достигает активного центра белка. Кроме того, фотоинактивация ЛДГ может быть связана с комплексированием молекул красителя с поверхностными участками белковой глобулы, что благоприятствует протеканию реакций по типу I, т.е. обусловлена непосредственным химическим взаимодействием триплетных состояний сенсибилизатора с аминокислотными остатками фермента.
Суммируя результаты исследования УФ-индуцированных изменений структурно-функционального состояния изоферментов ЛДГ в условиях их различного микроокружения, можно заключить, что основной тип УФ-модификации каталитической активности изоформ ЛДГ (мышечного и сердечного типов) — фотоинактивация ее молекул. На основании определения величин константы скорости инактивации и времени полупревращения изоферментов ЛДГ показано, что сердечная форма фермента (Н4) более фотостабильна по сравнению с мышечной (М4). Использование широкого спектра экзогенных соединений (азида натрия, 13- каротина, D-маннита, гистидина, серотонина, NADH), способных акцептировать АФК и проявлять в определенных концентрациях фотопротекторное действие по отношению к молекулам ЛДГ, позволило выявить существенную роль синглетного молекулярного кислорода в процессе фотоинактивации ЛДГ. При исследовании сенсибилизированного МГ фотоокисления изоформ
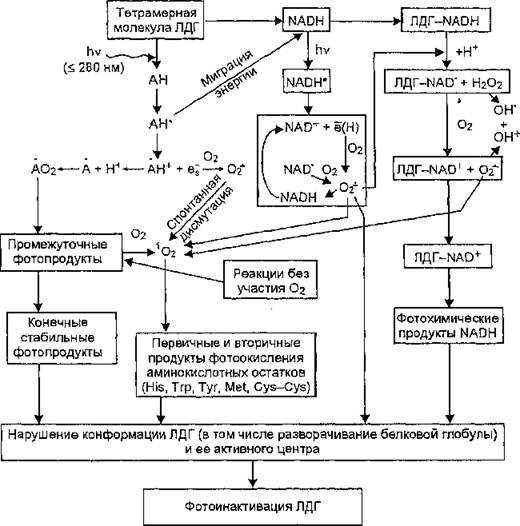
Рис. 59. Схема возможных физико-химических процессов, приводящих к фотоинактивации молекулы лактатдегидрогеназы
фермента показана возможность окисления ЛДГ (М4) синглетным кислородом при его экзогенной генерации.
На рис. 59 представлена схема возможных процессов, способных приводить к фотоинактивации ЛДГ.
Результаты проведенных исследований углубляют современные представления об особенностях фотохимических превращений и функциональных нарушений отдельных изоферментов ЛДГ, индуцированных воздействием УФ-излучения. Их необходимо учитывать при изучении первичных и начальных процессов УФ- изменений сложных белков с доменной структурой. Кинетические исследования фотоокисления ЛДГ в присутствии протекторов, акцептирующих активные промежуточные продукты фотопревращений белка, позволят оценить реальный вклад каждой элементарной стадии (реакции) в сложный процесс УФ-модификации субъединичной молекулы этого фермента.
КОНТРОЛЬНЫЕ ВОПРОСЫ
1. Какие компоненты биомембран являются акцепторами УФ-излучения?
2. Что понимают под термином “перокеидное фотоокисление липидов” мембран?
3. Каковы механизмы протекания процесса пероксидного фотоокисления липидов мембран?
4. Чем обусловлены изменения структурно-функционального состояния простых и сложных белков под влиянием УФ-излучения?
5. В чем заключаются особенности фотохимических превращений мембраносвязанных белков по сравнению со свободными?
6. Что представляет собой эффект фотохимической аллотопии? Каковы условия проявления и исчезновения феномена фотохимической аллотопии?
7. Что называют фотосенсибилизаторами?
S. Каковы механизмы протекания фотосенсибилизированных реакций?
9. В чем состоит роль синглетного молекулярного кислорода в реализации фотосенсибилизированных процессов?
10. Охарактеризуйте методы выявления и оценки роли синглетного кислорода в различных фотосенсибилизированных и темновых реакциях.
11. Что называют акцепторами и тушителями синглетного молекулярного кислорода? Как оценивают способность этих соединений взаимодействовать с ’О2?
12. Какие биологические молекулы являются наиболее эффективными тушителями синглетного кислорода, а какие — малоэффективными?
13. В чем заключаются различия химического и физического механизмов тушения Ю2 биомакромолекулами?
14. Каковы “последствия” фотосенсибилизированного повреждения молекулярных компонентов биомембран?
15. Опишите последовательность реакций, индуцированных воздействием УФ-света на отдельные мембранные компоненты и приводящих к нарушениям структурно-функционального состояния биомембран.
16. Какое теоретическое и практическое значение для медицины имеют исследования, направленные на изучение закономерностей фотохимических превращений биополимеров, их надмолекулярных комплексов и биомембран?