§ 7. Возникновение диффузионной неустойчивости в системе типа хищник — жертва
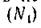
Пусть локальное взаимодействие между популяциями жертвы и хищника (N2) описывается системой двух обыкновенных дифференциальных уравнений
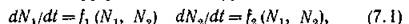
”—>рая имеет стационарное положительное решение
, Это решение устойчиво, если действительные части

Предположим, что параметры системы удовлетворяют неравенству, противоположному (7.6) (и, кроме того, удовлетворяют неравенствам (7.2)).
Тогда, какими бы ни были пространственные возмущения, локальные равновесия будут устойчивыми, и в пространстве будет сохраняться равно-11 Ю, М. Свирежев, Д. О. Логофет
мерное стационарное распределение численностей хищника и жертвы. Пусть теперь параметры системы медленно меняются, и в какой-то момент неравенство (7.6) стало выполняться при сохранении неравенств (7.2). Тогда, если в спектре пространственных возмущений присутствуют колебания с волновым числом &min, то их амплитуда начинает расти — развивается диффузионная неустойчивость, равномерность распределения нарушается и возникает пространственная периодическая структура. Дальнейшее развитие этого процесса может привести к развалу первоначально единого сообщества на отдельные, не связанные между собой ячейки.
В этом случае, хотя локальная мстойчивость равновесий и сохраняется, но ппи определяются
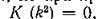
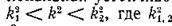
из уравнения .
. эти равновесия становятся диффузионно неустойчивыми.
Проиллюстрируем эти рассуждения на примере воль- терровской модели с
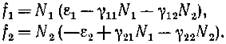
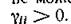
вида:
Поскольку „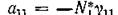 .и
.и условие (7.3)
условие (7.3)
че ипжрт быть выполнено НИ При КЗКИХ j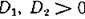 I, если
I, если
Поэтому мы рассмотрим модель несколько ИНОГО
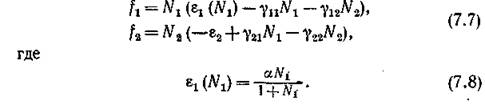
При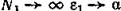 _ , и модель (7.7)—(7.8) совпадает с воль-
_ , и модель (7.7)—(7.8) совпадает с воль-
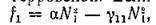
трпппвскпй. Если же Л\ мало, то (в отсутствие хищника) т. е. скорость размножения жертвы пропорциональна квадрату ее численности. Последнее хорошо описывает ситуацию двуполого размножения, когда рождение новых особей существенным образом зависит от вероятности образования репродуктивной пары.
Упростим модель, предположив, что смертностью за счет внутривидовой конкуренции у жертв и естественной смертностью хищника (сильная внутривидовая конкурен-
ция) можно пренебречь. Если, кроме того, ввести новые переменные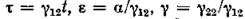 и положить, что
и положить, что
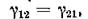 то уравнения локальной модели можно записать
то уравнения локальной модели можно записать
в виде

Из (7.11) видно, что если подвижность хищника намного превосходит подвижность жертвы то не
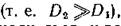
равенство (7.11) наверняка будет выполняться, и возникнет диффузионная неустойчивость.
Для более наглядного представления на рис. 45, а, б, в в плоскости параметров 6 =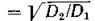 и є приведены графики кривых, разделяющих области диффузионной устойчивости и неустойчивости (для трех значений у). На рис. 46 приведена зависимость
и є приведены графики кривых, разделяющих области диффузионной устойчивости и неустойчивости (для трех значений у). На рис. 46 приведена зависимость от б (б пропорционально отношению радиусов индивидуальной активности хищника и жертвы).
от б (б пропорционально отношению радиусов индивидуальной активности хищника и жертвы). Из этих рисунков видно, что с ростом є — коэффициента рождаемости жертвы — для возникновения диффузионной неустойчивости подвижность хищника по сравнению с по
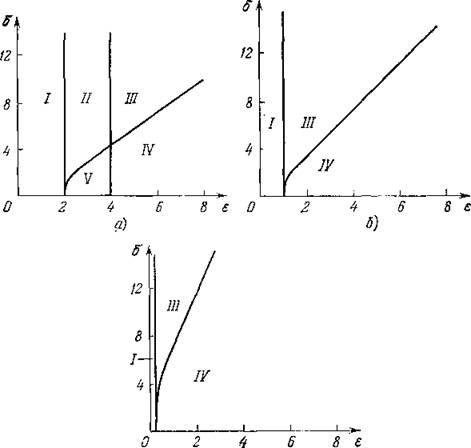
Рис. 45. Области различных типов устойчивости при нескольких значениях у: а) у = 0,5; б) у = 1; в) у = 5. Область / — в системе не существует нетривиального равновесия; II — равновесие существует, но оно неустойчиво как локально, так и диффузионно; /// — равновесие устойчиво локально, но неустойчиво диффузионно; IV — равновесие устойчиво как локально, так и диффузионно; V — равновесие локально неустойчиво, но устойчиво диффузионно.
6)
движностью жертвы должна также возрастать. Если же подвижность хищника мала, то диффузионная неустойчивость наверняка не развивается. Тот же самый эффект
наблюдается и с ростом у — коэффициента внутривидовой конкуренции среди хищников. Интересно, что при у