Схема развития генетического древа
Как и в случае с историей исхода из Африки, ответ на интересующий нас вопрос станет более ясным, если мы посмотрим на маркеры Адама и Евы. Большим достоинством построения древа митохондриевой ДНК и Y-хромосомы является то, что они позволяют проследить ветви и даже побеги, соответствующие отдельным молекулам, применительно к каждому конкретному району.
Эту увлекательную научную «игру» в гены и районы их распространения можно назвать филогеографией. B принципе она заключается в соединении разрозненных региональных побегов в крупные ветви и размещении последних на генетическом древе.Хотя правила предельно просты, их применение может оказаться достаточно сложным. Чтобы проследить векторы миграции из одного региона в другой, нам необходимо найти ветви-источники в обоих регионах и новый «побег» в искомом районе, который отсутствует на ветви прародины. Для прародины с многообразными векторами миграции в самых разных направлениях нам необходимо выявить общие ветви, имеющие разные «побеги» в других
Puc 5 5
районах. Пожалуй, один из наиболее удачных примеров подобной методики был описан в работе видного итальянского генетика Антонио Торрони и его коллег, опубликованной в 1993 г.
Эти исследователи выявили четыре основные линии митохондриевой ДНК этнической группы-родоначальницы коренного населения Америки, пометив их буквами А, В, C и D. Истоки каждой из этих буйно разросшихся ветвей, давших обильные «побеги» в Америке, можно возвести к аналогичным ветвям-прародителям А, В, C и D в Азии (см. главы 6 и 7).
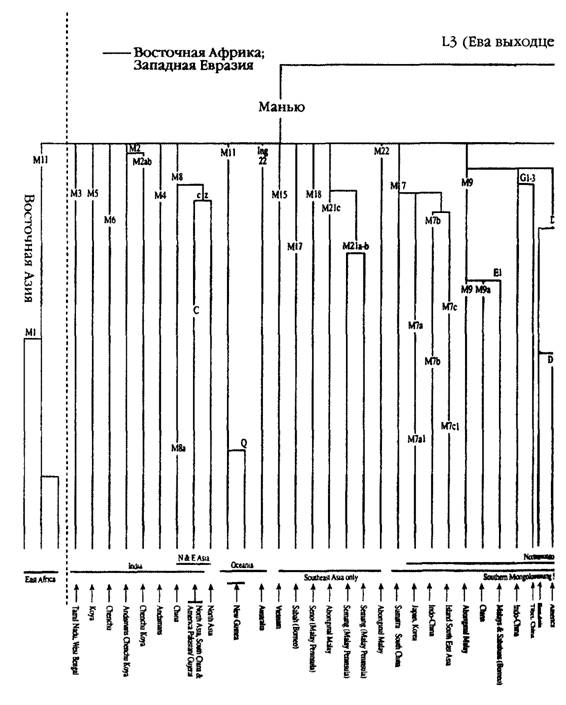
Мы можем применить этот же подход и к гораздо более сложной проблеме — идентификации прародины монголоидов в Азии. Взяв за начало митохондриевую ДНК Восточной Азии, мы можем заметить, что на сегодня там присутствуют девять четко сформировавшихся ветвей (га- плогрупп; см.
рис. 5.5), которые можно обозначить буквами A, D, С, D, E, F, G, X и Z. За исключением сестринских групп C и Z, ни одна из этих групп не может быть выведена на основе другой. Каждая из них более или менее уверенно выстраивается на основе двух евразийских линий- родоначальниц — Манью или Насрин. Недавно были описаны еще несколько второстепенных групп (M7/M10 и N9), однако их существование никак не сказывается на значимости аргументации. (Более подробно о фиголенезе и источниках по распределению гаплогрупп митохондриевой ДНК в Восточной Азии см. примечание03()Из девяти наиболее хорошо изученных восточно-азиатских ветвей митохондриевой ДНК три (С, X и Z, см. рис. 5.9) прослеживаются только к северу от условной линии «запад — восток», проведенной ниже Гималаев, тогда как остальные шесть (А, В, D, E, F и G) были обнаружены у обеих — как северной, так и южной — ветвей монголоидов. Ha первый вгляд может показаться, что эти уникальные северные ветви — весомые аргументы в пользу гипотезы O том, что прародина монголоидов находилась к северу от
Гималаев, но на самом деле у этого феномена есть более простое объяснение. Дело в том, что линия C и близкородственная ей линия Z, а также загадочная группа X не являются коренными фигурантами древнейшей истории Монголии. Они появились среди северной ветви монголоидов откуда-то извне, вероятнее всего — из Сибири или Центральной Азии. Как мы скоро увидим, в поддержку этой версии существует целый ряд аргументов. Распределение генов и структура шести остальных ветвей генетического древа свидетельствуют о том, что генетическая прародина монголоидов находилась к югу от Гималаев. C особенной ясностью на это указывают древнейшие генетические ветви B и F. Таким образом, мы, по всей видимости, имеем дело сразу с несколькими источниками генов у жителей Юго-Восточной Азии, которые впоследствии распространились на север. Наоборот, среди обитателей Северной Азии мы имеем несколько ветвей, никак не связанных с коренным населением Юго-Восточной Азии.
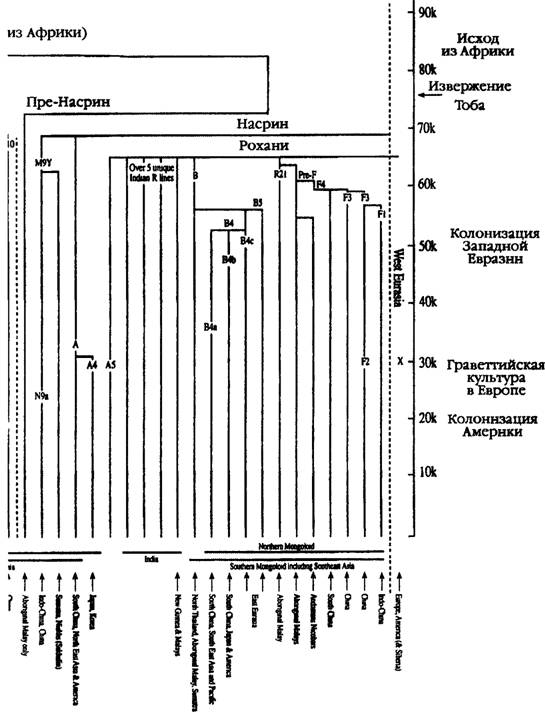
Возвращаясь к той же версии о развитии ветвей, можно отметить, что идея о южной прародине и последующей миграции монголоидов на север подтверждается данными исследования материнских генов и вполне соответствует гипотезе об эволюции сундадонтов в синодонтов (см. главу 5).Чтобы сосредоточить внимание на поисках возможного вторичного северного региона «дальнейшего развития той же темы», нам придется обратиться к обзору географических зон, где присутствуют все шесть ветвей мито- хондриевой ДНК — А, В, D, E, F и G, — являющихся общими для северной и южной ветвей монголидов. Таким образом, продвигаясь с запада на восток, мы обнаружим, что в Центральной Азии присутствуют пять основных линий (за исключением G), в Тибете — все шесть, в Монголии — пять (за исключением E), в Китае — четыре (за исключением E и G) и, наконец, в Корее — пять (за исключением Е)(34). Пожалуй, вряд ли можно считать простым совпадением, что страны с большим числом генетических линий, обычно ассоциируемых с монголоидной расой — Центральная Азия, Тибет, Монголия и Корея, — расположены вдоль 40-й параллели, которая, согласно Гатри, служила своего рода осью Мамонтовой степи (см. рис. 5.6).
Основываясь на этих факторах, мы можем задать вопрос о том, каков примерный возраст этих ветвей, расположенных вдоль 40-й параллели. Тоомас Кивисилд установил, что местный возраст четырех из этих ветвей в Монголии весьма близок: ветвь A насчитывает 35 500 лет, B — 40 500 лет или 33 500 лет (см. ниже), D — 44 500 лет и, наконец, F — 42 500 лет. Эти даты значительно «моложе», чем средний возраст тех же линий на юге, в Южном Китае. Ho самое важное заключается в том, что они вполне сопоставимы с датами начала активной колонизации Центральной Азии и относятся к эпохе после улучшения климата в Азии, которое произошло примерно 52 тысячи лет назад (см. главу 3) и благодаря которому первые обитатели Азии смогли проникнуть на просторы Мамонтовой сте- пи(35).
И все же: по каким путям и маршрутам люди современного типа проникли в этот обширный регион? Обра- тившсь к анализу двух древнейших линий Юго-Восточной Азии — B и F (идущих от потомков Насрина), которые тянутся из Юго-Восточной Азии в Тибет, Монголию и далее на северо-восток, вдоль побережья Тихого океана, мы видим, что они проникают в глубь материка в Сибири (Северная Азия), а затем останавливаются(36).
B этом отношении распределение этнических групп B и F отражает древнейшее распространение признаков сундадонтности. Как В, так и F достигают наиболее выраженной поливариантности среди туземных этнических групп Таиланда, Вьетнама и Камбоджи, что свидетельствует о том, что их прародиной является Индокитай.Весомым аргументом в поддержку версии о том, что группа F возникла и сформировалась на юге, является вновь идентифицированная гаплогруппа «пре-F», которая
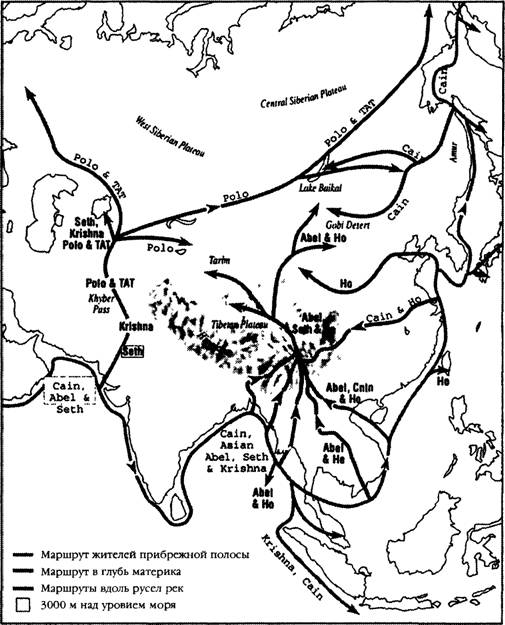
Рис. 5-6
распространена среди малайцев — коренных жителей Малайского полуострова (выводы сделаны на основе распространения признаков сундадонтности; см. рис. 5.6). Представители этой группы «пре-F» встречаются в Южном Китае (провинции Юннань и Гуаньси), Индокитае (Таиланд и Вьетнам), на Суматре, а также Андаманских и Никобарских островах, что, по всей вероятности, свидетельствует о древнейших этнических связях с обитателями прибрежной полосы в Юго-Восточной Азии(37).
Располагая сведениями о том, распространились ли представители группы B на запад, проникнув из Китая в Монголию вдоль Шелкового пути, или же двинулись на север из Бирмы через Тибет, мы сможем узнать направление маршрута первой волны колонизации, которая, по всей видимости, имела место около 50 тысяч лет тому назад. Это — немаловажные ключи. Тот же Тоомас Кивисилд установил, что возраст двух основных подгрупп В, проникших в Монголию, составляет 40 500 лет (Bl) и 33 500 лет (B2). Это говорит о том, что, хотя в Монголии и части западных районов Центральной Азии оба подтипа B могли появиться еще в эпоху палеолита, более древняя из этих ветвей могла пройти через Тибет, тогда как более поздняя, не исключено, проникла на запад, в Центральую Азию, с побережья Тихого океана, где ее представители сегодня занимают доминирующее положение(38).
Надо упомянуть и о трех линиях — D, E и G, — частично связанных с Манью, линиях, распространение представителей которых в Восточной Азии увеличивается при продвижении с севера на юг, отражая аналогичное распространение признаков синодонтности.
Однако остается нерешенным вопрос о том, выходцами откуда могли быть другие основатели «этнической КОЛОНИИ МОНГОЛОИДОВ» B Мамонтовой степи (С, X и Z) и каким именно из трех возможных коридоров, ведущих в Мамонтовую степь, они воспользовались. K югу от Гималаев представители этих трех линий Северной Евразии встречаются крайне редко. Ветви-сестры C и Z наиболее распространены в Сибири и Северо-Восточной Азии, тогда как в более южных районах (за исключением Монголии, Тибета и Центральной Азии) их представители почти не встречаются. Линии эти тянутся вдоль всего континуума «восток — запад» через азиатские великие степи, и численность их представителей вЦентральной Азии вплоть до Турции весьма значительна. Что касается ветви С, то она простирается даже в Америку через Аляску, однако преимущественное ее распространение на севере Восточной Азии свидетельствует о том, что эти люди не могли прийти в Центральную Азию через Китай. Наоборот, куда более вероятно, что они проникли в Азию откуда-то с запада, причем представители восточной ее ветви были обладателями новой технологии создания орудий, характерной для Позднего палеолита, артефакты которой были обнаружены в пещере Кара-Бом на Алтае (Россия). Возраст их — около 43 тысяч лет. Люди, создавшие их, принадлежали к группе Манью, и их древнейшие предки пришли сюда из Индии и Пакистана, проникнув в долину Инда через Кашмир или Афганистан и обогнув по пути западную оконечность Гималаев (см. рис. 5.4). Bce эти факты свидетельствуют о том, что мигранты проникли в азиатскую степь где-то 40—50 тысяч лет тому назад из западных районов Центральной Азии (см. рис. 5.6). Подсчеты Тоомаса Кивисилда, находящиеся в полном соответствии с версией о том, что линии C и Z распространялись по степи и их носителями были обладатели новой технологии эпохи Верхнего палеолита, показывают, что возраст ' группы C в Монголии составляет 42 тысячи лет(39).
Последняя из северных линий, линия X, обнаружена только у европейцев и туземных жителей Америки (правда, есть сообщение о находке следов линии X в Южной Сибири), однако связь между Старым и Новым Светом существовала еще как минимум 30 тысяч лет назад.
Это свидетельствует о том, что группа X распространялась по необъятным пространствам Центральной и Северо-Восточной Азии вместе с представителями культуры охотников на мамонтов, однако полностью исчезла в Азии во время последнего Ледникового периода, сохранившись лишь на отдаленных окраинах своего ареала в Америке и Европе (см. главу 7)(40).Столь сложная картина линий распространения мито- хондриевой ДНК свидетельствует O том, что монголоиды были выходцами с юга, тогда как жители Центральной Азии происходят от некоей западно-азиатской этнической группы, однако в формирование их родовых признаков внесли свой вклад уроженцы Восточной и Юго-Восточной Азии, мигрировавшие в Центральную и Северо- Восточную Азию. Эти генетические данные являются аргументами в поддержку теории о трех этапах колонизации Центральной Азии с «баз» на побережье Индийского и Тихого океанов 40—50 тысяч лет тому назад. Разграничение и выделение генетических составляющих в этносах и этнических группах Сев