Динамика растительной массы и биологический круговорот.
По величине годичного прироста (13—38 ц/гатод) экосистемы кедрового стланика (рис. 2) относятся к очень малопродуктивным и малопродуктивным природным комплексам (Родин, Базилевич, 1965).
В зависимости от экологических условий и возраста растений участие основных компонентов фитомассы в создании прироста варьирует в широких пределах: фотосинтезирующих органов от 48 до 64 %, многолетних одревесневших частей от 7 до 24 % и корней от 20 до 33 % от величины общего прироста. Особенностью характеризуемых экосистем является то, что более половины прироста приходится на кедровый стланик. Это существенно отличает кедрово-стланиковые заросли от ландшафтов лесотундры, в которых динамика растительной массы обусловлена в основном напочвенным покровом (Родин, Базилевич, 1965).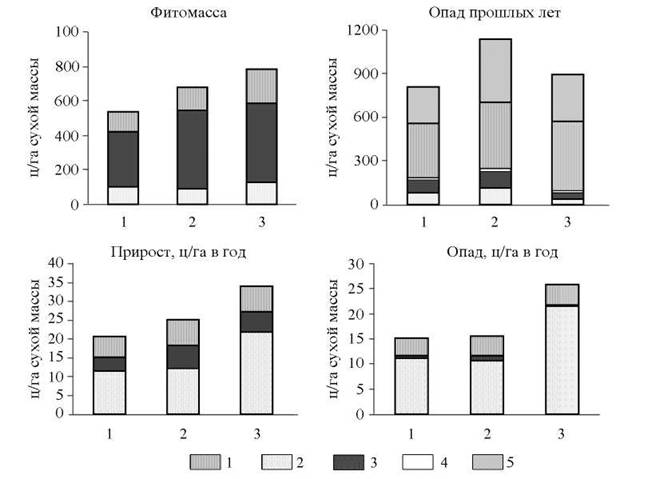
Рис. 2. Показатели динамики растительной массы в экосистемах (1 - шикшево-лишайниковых, 2 - багульниковобрусничных, 3 - шикшево-долгомошных) кедровых стлаников, ц/га сухой массы: 1 - корни; 2 - фотосинтезирующие органы; 3 - многолетние органы; 4 - мхи и лишайники; 5 — измельченный опад корней
Прирост фитомассы рассматриваемых экосистем близок к приросту тундр Восточно-Европейского Севера (Данилов, 1958), кустарничковых тундр Хибинских гор (Чепурко, 1972), он составляет около половины годового прироста северо-таежных лесов Архангельской области (Руднева и др., 1966). Следовательно, по величине и структуре годичного прироста кедрово-стланиковые заросли близки к типичным тундровым сообществам.
Ежегодный опад составляет 1/30-1/42 часть всей фитомассы. В его составе доминируют фотосинтезирующие органы (66-83 %) и корни (16—31 %); роль стволовой древесины и ветвей невелика: 1-14 %. Таким образом, по общим показателям прироста и опада экосистемы кедрового стланика в целом можно отнести к типичным тундровым комплексам.
Однако, учитывая их свое- образне, следует согласиться с мнением И.А. Соколова (1973) о целесообразности выделения пояса стлаников в качестве самостоятельной географической зоны субполярных областей Восточной Сибири и Дальнего Востока.В зависимости от экологических условий и структуры биомассы роль отдельных растений в формировании опада различна: в кедровнике шикшево-лишайниковом существенно участие лишайников, а в кедровнике шикшево-долгомошном - осок.
В составе опада кедрового стланика доминирует хвоя - 85-90 %. С увеличением возраста экосистем участие хвои в опаде относительно уменьшается и возрастает роль почечных чешуек (4,5-7,6 %), коры, пыльников (0,1-3,7 %) и шишек (0,0-7,8 %). Хвоя кедрового стланика опадает постепенно в течение всего вегетационного периода; максимум опада отмечается в конце августа- середине сентября.
По массе ежегодного опада кедрово-стланиковые заросли вполне сопоставимы с низкобони- тетными и заболоченными лесами хвойно-таежной зоны. Бедность растительных остатков зольными элементами и азотом, кислая реакция опада и среды разложения, длительное пребывание почв в мерзлом состоянии, малочисленность и малая активность микрофлоры, неблагоприятное сочетание гидротермических условий, богатство хвои восками и смолами не благоприятствуют деструкции отмерших частей растений (Берман и др., 1979).
Изучение скорости разложения опадов показало, что низкая влажность подстилок и антибиотические свойства лишайников обусловливают медленные темпы минерализации хвои в кедровнике шикшево-лишайниковом (табл. 8). Увеличение влажности органогенных горизонтов и уменьшение в составе напочвенного покрова лишайников способствуют более высоким темпам разложения опада хвои в кедровнике багульниково-брусничном и шикшево-долгомошном. Наиболее интенсивно разлагается опад листьев рододендрона, далее следуют листья осок, брусники и березы Миддендорфа. Наименее активно разлагаются отмершие части мхов.
Сопоставление активности разложения опадов различных растений с их химическим составом свидетельствует о наличии корреляции с содержанием в них азота и кальция.
Взяв за основу темпы деструкции опадов различных растений в первые 3 года, рассчитали ориентировочный срок их полной деструкции. Установлено, что для полного разложения годичного опада мхов требуется не менее 16 лет, хвои кедрового стланика - 8-14, листьев голубики, березы, брусники и багульника - 7-9, листьев рододендрона и осок - 5-6 лет.Сравнительно быстрые темпы разложения листьев голубики, брусники, багульника, березы и рододендрона в какой-то мере компенсируют степень участия высвобождающихся при их деструкции химических элементов в биологическом круговороте, несмотря на незначительное участие этих видов растений в создании массы годичного опада.
Несоответствие между относительно большим поступлением отмерших частей растений на поверхность почвы и низкими темпами их разложения реализуется в формировании грубогумусовых подстилок и обогащении мине-
Таблица 8. Темпы разложения опада доминантных видов растений ральной части ПОЧВЄИИОГО профиля экосистем кедрового стланика зоны Тауйской губы, % от первона- ИЗМельченным корневым опадом чальнои массы нарастающим итогом
| Вид опада | Период разложения, год | |||
| 1-й | 2-й | 3-й | В среднем | |
| Хвоя кедрового стланика | 13,9 | 29,2 | 35,6 | 11,9 |
| Листья березы Миддендорфа | 23,8 | 33,9 | 38,0 | 12,7 |
| Листья рододендрона | 11,0 | 23,6 | 63,0 | 21,0 |
| Листья багульника болотного | 22,2 | 30,3 | - | 15,1 |
| Листья брусники | 22,2 | 29,7 | 41,2 | 13,7 |
| Листья голубики | 17,7 | 27,2 | 34,6 | 16,5 |
| Листья осоки круглой | 26,2 | 38,0 | 47,2 | 15,7 |
| Отмершие части политриха | 4,7 | - | 19,3 | 6,4 |
разной степени гумификации, основную массу которого составляют корни кедрового стланика и кустарничков; на долю отмерших надземных органов растений приходится 10- 27 % от общего запаса мертвой массы.
Превышение опада прошлых лет над массой ежегодно отмирающего вещества в зависимости от экологических условий местообитания варьирует от 34 в кедровнике шикшево- долгомошном до 77 в кедровнике ба-Примечая и е. Прочерк - данные не определялись. гульниково-брусничном. Это дает основание рассматривать кедрово-стланиковые заросли как ландшафты с застойным или сильно заторможенным типом биологического круговорота веществ (по: Родин, Базилевич, 1965).