Биологический круговорот.
Величины ежегодного потребления химических элементов приростом растительной массы в ландшафтах кедрового стланика невелики (рис. 3): 44- 104кг/га-год , в том числе на фотосинтезирующие органы приходится 68-82 %, многолетние одревесневшие части 5-18 % и корни 11-23 % от общего поглощения.
Подавляющее количество зольных элементов и азота аккумулируется хвоей кедрового стланика; в кедровнике шикшево-дол- гомошном велика также и роль осок; менее активно накапливаются элементы-органогены приростом стволов, ветвей и корней стланика. Участие кустарничков в этом процессе минимально.Во всех исследованных фитоценозах наиболее интенсивно аккумулируется азот. В кедровниках шикшево-лишайниковом, багульниково-брусничном и багульниково-лишайниковом второе место по этому показателю занимает группа элементов, включающая кальций и калий; в кедровнике шикшево-долгомошном - кремний, калий и кальций.
Косвенная характеристика интенсивности аккумуляции химических элементов из почвы приростом, полученная путем вычисления рядов биологического поглощения, показала, что во всех кедрово-стланиковых ландшафтах из минеральной части почвенных профилей наиболее энергично поглощаются кальций, затем калий и марганец.
Таблица 9. Содержание азота и зольных элементов в составе доминантных видов растений экосистем кедрового стланика, % к сухому веществу
| Растения и их части | Чистая зола, % | N | Si | Са | К | Mg | Р | А1 | Fe | Мп | S | Na |
| Кедровый | стланик | |||||||||||
| Хвоя 1-го года | 2,91 | 1,36 | 0,20 | 0,74 | 0,54 | 0,18 | 0,05 | 0,09 | 0,14 | 0,03 | 0,13 | 0,01 |
| прошлых лет | 2,16 | 1,31 | 0,16 | 0,46 | 0,34 | 0,22 | 0,03 | 0,11 | 0,08 | 0,14 | 0,14 | СЛ. |
| Ветви 1-го года | 2,08 | 1,34 | 0,02 | 0,36 | 0,68 | 0,19 | 0,06 | 0,04 | 0,04 | 0,03 | 0,19 | 0,02 |
| прошлых лет | 0,98 | 0,43 | 0,05 | 0,20 | 0,23 | 0,06 | 0,04 | 0,07 | 0,02 | 0,03 | 0,15 | СЛ. |
| ствол | 0,34 | 0,11 | 0,03 | 0,09 | 0,04 | 0,02 | 0,01 | 0,01 | 0,01 | СЛ. | 0,10 | сл. |
| Корни 0,7-0,5 см | 0,61 | 0,17 | 0,03 | 0,22 | 0,04 | 0,04 | 0,02 | 0,03 | 0,01 | 0,01 | 0,08 | сл. |
| менее 0,7 см | 0,96 | 0,51 | 0,03 | 0,19 | 0,20 | 0,06 | 0,02 | 0,03 | 0,04 | 0,01 | 0,09 | 0,01 |
| Береза Миддендорі | >а | |||||||||||
| Листья | 2,86 | 1,41 | 0,08 | 0,91 | 0,21 | 0,22 | 0,11 | 0,07 | 0,10 | 0,16 | 0,09 | 0,01 |
| Ветви | 1,00 | 0,56 | 0,04 | 0,28 | 0,15 | 0,07 | 0,03 | 0,02 | 0,04 | 0,02 | - | сл. |
| Корни | 1Д1 | 0,39 | 0,04 | 0,30 | 0,17 | 0,08 | 0,04 | 0,03 | 0,02 | 0,03 | 0,05 | 0,01 |
| Рябина бузинолистная | ||||||||||||
| Листья | 5,07 | 2,05 | 2,02 | 1,17 | 1,34 | 0,27 | 0,33 | 0,01 | 0,01 | 0,07 | 0,14 | 0,02 |
| Ветви | 2,14 | 0,43 | 0,01 | 0,43 | 0,02 | 0,11 | 0,09 | 0,01 | - | 0,03 | 0,01 | 0,02 |
| Рододендрон золотистый | ||||||||||||
| Листья | 3,09 | 0,78 | 0,03 | 0,86 | 0,25 | 0,61 | 0,06 | 0,10 | 0,03 | 0,04 | - | 0,03 |
| Стебли | 1,59 | 0,53 | 0,06 | 0,31 | 0,19 | 0,12 | 0,04 | 0,08 | 0,04 | 0,18 | 0,07 | 0,01 |
| Корни | 0,32 | 0,40 | 0,03 | 0,09 | 0,02 | 0,02 | 0,01 | 0,01 | СЛ. | 0,01 | 0,12 | 0,01 |
| Брусника | ||||||||||||
| Листья | 2,22 | 0,60 | 0,10 | 0,64 | 0,26 | 0,11 | 0,10 | 0,02 | 0,12 | 0,10 | 0,16 | 0,02 |
| Стебли | 2,36 | 0,56 | 0,07 | 0,59 | 0,27 | 0,12 | 0,06 | 0,15 | 0,10 | 0,16 | - | 0,02 |
| Корни | 0,42 | 0,52 | 0,01 | 0,11 | 0,07 | 0,04 | 0,01 | 0,01 | 0,01 | 0,03 | - | 0,01 |
| Голубика обыкновенная | ||||||||||||
| Листья | 2,86 | 1,30 | 0,16 | 0,60 | 0,55 | 0,55 | 0,11 | 0,03 | 0,08 | 0,06 | - | 0,01 |
| Стебли | 1,34 | 0,41 | 0,03 | 0,29 | 0,26 | 0,04 | 0,04 | 0,07 | 0,04 | 0,15 | - | 0,01 |
| Багульник стелющийся | ||||||||||||
| Листья | 2,51 | 1,44 | 0,18 | 0,70 | 0,47 | 0,12 | 0,07 | 0,01 | 0,06 | 0,05 | 0,12 | 0,02 |
| Стебли | 1,05 | 0,58 | 0,05 | 0,22 | 0,12 | 0,08 | 0,03 | 0,03 | 0,04 | 0,04 | - | 0,01 |
| Корни | 0,54 | 0,37 | 0,02 | 0,12 | 0,07 | 0,05 | 0,02 | 0,02 | СЛ. | 0,05 | - | СЛ. |
| Шикша черная | ||||||||||||
| Листья | 3,08 | 0,84 | 0,04 | 1,07 | 0,46 | 0,23 | 0,04 | 0,02 | 0,14 | 0,10 | 0,16 | 0,02 |
| Стебли | 1,32 | 0,51 | 0,09 | 0,31 | 0,15 | 0,11 | 0,03 | 0,04 | 0,06 | 0,05 | 0,04 | 0,01 |
| Корни | 0,72 | 0,33 | 0,05 | 0,15 | 0,12 | 0,04 | 0,03 | 0,03 | 0,01 | 0,04 | - | СЛ. |
| Филлодоца | ||||||||||||
| Листья | 2,56 | 1,02 | 0,03 | 0,90 | 0,20 | 0,23 | 0,08 | 0,02 | 0,16 | 0,01 | - | 0,02 |
| Стебли | 1,43 | 0,49 | 0,10 | 0,31 | 0,19 | 0,08 | 0,06 | 0,05 | 0,04 | 0,08 | - | 0,01 |
| Корни | 0,83 | 0,27 | 0,03 | 0,14 | 0,11 | 0,05 | 0,03 | 0,04 | 0,02 | 0,06 | - | 0,01 |
| Политрих обыкновенный | ||||||||||||
| Зеленые части | 1,50 | 0,91 | 0,25 | 0,30 | 0,20 | 0,04 | 0,03 | 0,04 | 0,06 | 0,02 | - | 0,10 |
| Бурые части | 2,65 | 0,51 | 0,42 | 0,67 | 0,18 | 0,04 | 0,07 | 0,06 | 0,12 | 0,04 | - | 0,02 |
| Дикран | ||||||||||||
| Зеленые части | 2,87 | 0,85 | 0,27 | 0,59 | 0,35 | 0,36 | 0,09 | 0,06 | 0,05 | 0,03 | 0,15 | 0,03 |
| Бурые части | 5,70 | 0,45 | 0,38 | 1,30 | 0,83 | 0,42 | 0,10 | 0,21 | 0,18 | 0,07 | 0,21 | 0,15 |
| Кладония оленья | 1,26 | 0,22 | 0,16 | 0,26 | 0,09 | 0,07 | 0,05 | 0,02 | 0,11 | 0,01 | 0,07 | 0,02 |
П р и м е ч а и и е. Прочерк - данные не определялись.
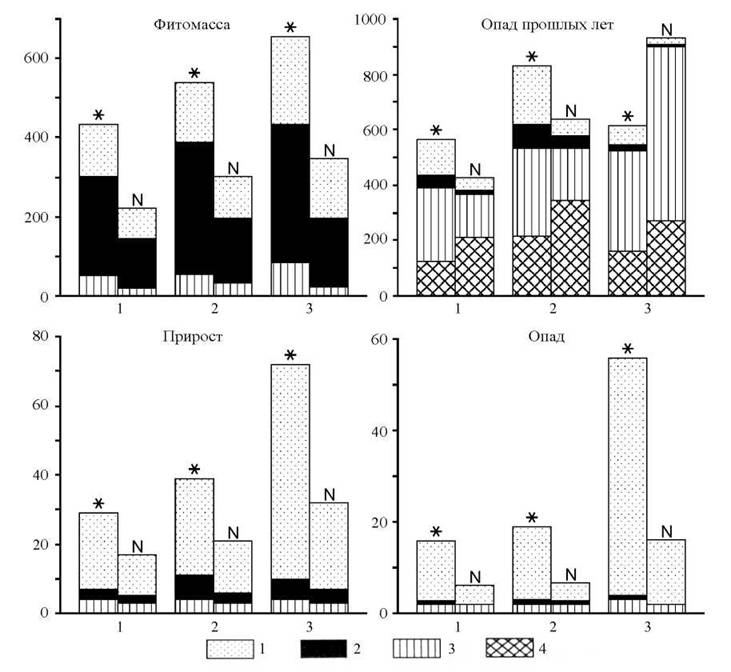
Рис. 3. Показатели биологического круговорота зольных элементов и азота в экосистемах шикшево-лишайникового (1), багульниково-брусничного (2), шикшево-долгомошного (3) кедрового стланика, кг/га; 1 - фотосинтезирующие органы, 2 - многолетние органы, 3 - корни, 4 - измельченный опад корней; звездочка - зольные элементы; N - азот
Обращает на себя внимание активное потребление железа в кедровниках шикшево- лишайниковом и багульниково-брусничном, с чем, видимо, связано относительное накопление его в верхних минеральных горизонтах; в кедровнике шикшево-долгомошном коэффициент поглощения Fe меньше единицы, что коррелирует с интенсивной его аккумуляцией в иллювиальной части профиля (Игнатенко и др., 1977).
Снижение степени элювиальности ландшафтов обусловливает уменьшение активности поглощения биофильных элементов. В отличие от лесотундры и тайги Кольского полуострова, кед- рово-стланиковые заросли характеризуются меньшей избирательной способностью по отношению к марганцу и калию и большей - кальцию и железу (Манаков, 1970, 1972).
Учитывая активное вовлечение кальция в биологический круговорот, можно предположить, что неусредненность кислого органического вещества в почвенном профиле подзолов и подбуров обусловлена медленным высвобождением щелочно-земельных элементов из разлагающегося опа- да и быстрым вовлечением их в биологический круговорот.
Накопление химических элементов в фитомассе экосистем стланика варьирует от 546 до 1000 кг/га, в том числе на фотосинтезирующие органы приходится от 19 до 37 %, многолетние одревесневшие части - 53-60 % и корни - 10-28 % (см. рис. 3). Значительное накопление химических элементов в фотосинтезирующих органах кедровника багульниково-лишайникового (52 %) обусловлено значительной охвоенностью ветвей и развитием кустарничково-лишайникового покрова.
По содержанию химических элементов кедрово-стланиковые заросли близки с низкобони- тетными лесами и болотами Европейской части России.
Из общей суммы на долю азота приходится до 40 %. Вторую группу элементов по активности их накопления в фитомассе образуют кальций, калий и сера. Такой характер аккумуляции, за исключением серы, присущ ельникам таежной зоны, некоторым сообществам восточно-европейской лесотундры и кустарничково-лишайниковым тундрам Кольско-карельской провинции (Марченко, Карлов, 1962; Руднева и др., 1966; Манаков, 1970; Игнатенко и др., 1973).По сравнению с приростом в фитомассе менее активно накапливаются магний, железо, фосфор, алюминий, марганец и, отчасти, сера, что связано с изменением химического состава растений с возрастом.
Роль зеленых фотосинтезирующих органов в аккумуляции химических элементов возрастает с 11-12 % от участия их в общей биомассе до 23-29 % в сумме минеральных веществ. Это несколько выше, чем в ельниках-зеленомошниках крайней северной тайги (Марченко, Карлов, 1962). Больше половины общего запаса элементов аккумулируется многолетними надземными органами. Следовательно, распределение химических элементов в фитомассе рассматриваемых экосистем аналогично распределению их в редкостойных, низкобонитетных зеленомошных лесах лесотундровой и таежной зон европейской части России. Однако аккумуляция в них азота более интенсивна.
Количество ежегодно возвращаемых с опадом химических элементов значительно ниже их ежегодного потребления: 20-71 кг/га, в том числе на долю фотосинтезирующих органов приходится 66-93 %, многолетних надземных частей - 1-7 % и корней - 7-27 %. Подавляющая часть химических элементов сосредоточена в хвое кедрового стланика. Доля участия в составе опада листьев кустарничков, лишайников и мхов значительно меньше и существенно различается в сравниваемых фитоценозах.
Средневзвешенное содержание в опаде азота составляет около 0,6 %, т. е. является чрезвычайно низким по сравнению с фитоценозами более южных широт. Оно возрастает от кедровников горных склонов (1,00-1,03 %) к кедровникам межгорных долин (2,11 %), характеризующимся развитым травяно-моховым покровом.
Экосистемы кедрового стланика заметно различаются по соотношению возвращаемых химических элементов. В кедровниках шикшево-лишайниковом и багульниково-брусничном наиболее активно возвращаются азот и кальций, в кедровнике шикшево-долгомошном - кремний, азот и кальций. По сравнению с приростом, активность возврата кальция, калия, кремнезема и марганца выше, чем их ежегодное потребление; фосфора, серы, натрия, наоборот, ниже.
Значительная часть возвращаемых химических элементов накапливается в опаде прошлых лет. Их количество в кедровнике багульниково-брусничном превышает ежегодный опад в 65 раз, в шикшево-лишайниковом - 48 раз, шикшево-долгомошном - 46 раз. Ряды накопления элементов в рассматриваемом компоненте биомассы следующие (в порядке убывания): в шикшево-бруснично- лишайниковом - кальций, азот, калий, магний, кремний, алюминий, железо, сера, фосфор, натрий; багульниково-брусничном - кальций, азот, кремний, калий, магний, алюминий, марганец, железо, сера, фосфор, натрий; шикшево-долгомошном - кремний, азот, кальций, калий, магний, фосфор, алюминий, марганец, железо, сера, натрий. Сравнение этих рядов с рядами по приросту показывает, что активность возврата кальция, калия, кремния и марганца выше, чем их поглощение, а фосфора, серы и натрия - ниже.
При сопоставлении характера консервации химических элементов в подстилках двух контрастных типов кедровников выявляется следующее: с переходом от кедровника шикшево- лишайникового к кедровнику багульниково-брусничному относительно увеличивается накопление азота, кальция, калия, марганца и уменьшается - кремния, алюминия, серы, магния. Это обусловлено как геохимическими особенностями сравниваемых ландшафтов, так и изменениями структуры подстилок.
Под действием процессов минерализации и гумификации химический состав мертвых растительных остатков претерпевает значительные изменения. Анализ рядов выноса-накопления указывает на то, что в опаде растительной массы прошлых лет по сравнению со свежим опадом происходит активное накопление серы, железа и азота.
Такие элементы, как кремний, кальций, калий, магний, алюминий и марганец, в одних случаях выносятся, в других - аккумулируются. На более поздних стадиях разложения происходит интенсивное накопление кремния, кальция, магния и азота. Значительно слабее выражена аккумуляция фосфора, железа и алюминия. Содержание калия и марганца в торфянистых горизонтах заметно снижается, что связано как с вовлечением их в новый цикл биологического круговорота, так и с выносом в минеральные горизонты почвы.Следовательно, основные биофильные элементы (азот, кальций, возможно, калий) энергично накапливаются в отмершей растительной массе и лишь отчасти выносятся за пределы ландшафта. В подземной части кедрово-стланиковых фитоценозов сосредоточено от 47 до 72 % от общих запасов зольных элементов и азота. Основная масса их аккумулируется в сухоторфянистых и перегнойных горизонтах. Существенное участие в консервации химических элементов принимают измельченное органическое вещество и мелкие корни, среди которых преобладают отмершие.
По качественному составу основных аккумулируемых элементов надземные и подземные части экосистем близки: азот, кальций, калий. Особенностью является увеличение участия железа, кремния и алюминия в корневой массе, что обусловлено наличием трудно отделимых механических примесей и пропитыванием опада минерализованными почвенными растворами.
Основными химическими элементами, накапливающимися в общей биомассе кедрово- стланиковых ландшафтов, являются азот, кальций и калий. Значительно менее интенсивно аккумулируются остальные зольные элементы. В порядке убывания они располагаются в следующий ряд: железо, кремний, сера, магний, алюминий, фосфор, марганец, натрий. Запасы железа превышают таковые кремния в фитоценозах со значительным участием лишайников; магний, сера и алюминий не обнаруживают четких закономерностей; фосфор, марганец и натрий накапливаются в минимальных количествах.
Контрастность биологического круговорота, представляющая собой отношение содержания химических элементов в составе биомассы аккумулятивно-элювиального элементарного ландшафта к емкости смежного элювиального ландшафта, находящегося в геохимическом сопряжении с первым, возрастает при переходе от северотаежных биогеоценозов (0,84 : 1,00) к кедрово-стла- никовым (0,77 : 1,00). В соответствии с классификацией Л.Е. Родина и Н.И. Базилевич (1965) кедрово-стланиковые ландшафты могут быть охарактеризованы как (калиево-) кальциево-азотные, низкозольные, очень мало- и малопродуктивные, застойные и сильнозаторможенные.