Глава 7. БИОЛОГИЧЕСКОЕ ДЕЙСТВИЕ ЛАЗЕРНОГО ИЗЛУЧЕНИЯ
Сам факт мощного (и управляемого) химического действия лазерного излучения, которое мы рассматривали выше, вызывает вопрос о практическом использовании такого излучения в медикобиологических задачах.
В настоящее время именно биологическое действие лазерного излучения представляет наибольший практический интерес и, вместе с тем, относится к наименее ясным вопросам лазерной физики. Парадоксально, но эта неясность связана не с недостатком результатов, а, напротив, с огромным количеством разнообразных экспериментальных данных. При этом наиболее загадочным оказалось действие низкоинтенсивного лазерного излучения, которое подробнее мы рассмотрим в данной главе.Видимо, следует оставить в стороне примитивные и часто совсем шарлатанские интерпретации биологического действия лазерного излучения типа «энергоинформационного поля», «биополя» и др., которые не имеют смысла уже потому, что пока не предсказали ни одного эффекта. Нельзя сказать, что эти объяснения противоречивы. Более того, такие объяснения всегда «правильны», поскольку, за счет смутных (умозрительных) определений невероятно большого числа основных понятий, они в принципе не могут содержать никаких противоречий. Как следствие, эти модели всякий раз легко модифицируются при объяснении новых наблюдений. Ясно, что все это является просто формой средневековой схоластики и не может иметь отношения к серьезной науке, которая всегда имеет целью давать проверяемые предсказания, а не просто объяснения.
Невозможно рассматривать методы анализа процессов в каком- либо объекте, не ознакомившись, хотя бы кратко, с основными свойствами этого объекта. Поэтому для анализа биологического действия лазерного излучения приведем некоторые сведения из биологии и биофизики. Эти сведения, конечно, ни в коей мере не составляют содержание биофизики, а лишь акцентируют внимание на некоторых (важных для дальнейшего) особенностях живых организмов.
Некоторые сведения из биофизики. Строение биологических
объектов. Прежде всего необходимо представлять себе строение
биологических объектов. Основным - всегда имеющимся - элементом биообъектов является клетка (см. рис. 7.1). Клетка ограничена в пространстве клеточной мембраной, представляющей собой двойной слой молекул фосфолипидов. Фосфолипид, как ясно из названия, это молекула, состоящая из липидного (аналогичного жирам, и следовательно, гидрофобного) и фосфатного (гидрофильного) остатков. Такие, как говорят, амфифильные молекулы, оказавшись в воде стремятся упаковаться таким образом, чтобы «экспонировать» в воду свои гидрофильные остатки и максимально «скрыть» от воды гидрофобные. Такие упаковки, естественно, просто минимизируют свободную энергию системы. Это приводит ко множеству вариантов упаковок, одной из которых и является двойной слой: гидрофильные остатки верхнего слоя и нижнего направлены наружу, а между ними «скрыты» гидрофобные липидные остатки. Часто, имея в виду клеточную мембрану, говорят просто «билипидный слой».
Обмен веществом с внешней средой в клетке осуществляется за счет мембранных белков («машин», в общем случае выполняющих довольно сложные функции). Так, трансмембранные белки образуют т.н. ионные каналы, через которые проходят ионы определенного сорта в зависимости от состояния белка.
Внутри клетки содержатся органеллы: ядро, рибосомы, митохондрии и, в зависимости от клетки, другие органеллы (комплекс Гольджи и т.п.). Каждая органелла также окружена либо мембраной, либо, чаще, белковой оболочкой. В ядре находится основной генетический материал клетки: молекулы ДНК (дезоксирибонуклеиновой кислоты), зафиксированные в т. н. хромосомах с помощью белков-гистонов. Все молекулы ДНК представляют собой линейные (в спиральной конформации) полимерные макромолекулы, образованные всего четырьмя мономерными элементами (нуклеотидами): аденином (А), цитозином (C), гуанином (G) и тимином (Т). Информация закодирована в последовательности нуклеотидов.
Основными исполнительными элементами клетки являются белки. Синтез белков, необходимых клетке, происходит в рибосомах по информации, считанной в ДНК и перенесенной к рибосоме с помощью транспортного РНК (рибонуклеиновой кислотой, аналог ДНК, в котором нуклеотид тимин заменен урацилом). Все белки состоят из всего 20 аминокислот (точнее, 27 с учетом растительных клеток). Известно, что каждая аминокислота белка в ДНК кодируется тройкой (т.н. «триплетом») нуклеотидов. Белки представляю собой линейные полимерные макромолекулы, в которой мономеры - аминокислоты - связаны так называемой полипептидной цепью. Последовательность аминокислот называется первичной структурой белка. В пространстве эта цепь упакована обычно сложным, неупорядоченным, но вовсе не случайным (т.е. вполне закономерным) образом. Пространственная структура макромолекулы называется ее конформацией. Для каждого белка существует огромное количество возможных конформаций. «Осмысленные» функции (например, ферментативный катализ) белки осуществляют только в какой-то определенной конформации, называемой нативной. Конформации относят к вторичной структуре белка. Как теперь известно, первичная структура белка полностью определяет его нативную конформацию. Различают также более высокие уровни организации белков: так называемые третичные и четвертичные структуры.
Основные химические процессы в клетках осуществляются с помощью белков в т.н. реакциях ферментативного катализа, где белки играют роль высокоспецифичных катализаторов. Молекула исходного вещества - субстрата - «захватывается» определенным конформационным элементом белка-фермента - так называемым активным центром. Белок, претерпевая последовательность кон- формационных переходов, вызванных неравновесностью нового состояния (белок + субстрат), превращает субстрат в продукт, который отщепляется от белка. Обратными конформационными переходами белок возвращается в нативную конформацию.
Для осуществления этих и большинства других процессов в клетке требуются затраты энергии.
Основной «энергетической» молекулой является аденозинтрифосфорная кислота (АТФ). Эта молекула выделяет энергию при расщеплении до АДФ и АМФ (аденозиндифосфорная и аденозинмонофосфорная кислота). Накопление энергии - синтез АТФ за счет приходящих реагентов происходит в митохондриях.Заметим, что все это - только общая грубая схема клетки. В действительности клетки могут сильно различаться по строению.
Так, например, характерные размеры большинства клеток - от 1 до 10 мкм, однако, клетки нервной ткани - нейроны могут иметь отростки - аксоны длиной около метра; существуют клетки, не имеющие ядра (например, эритроциты) и т.п.
Вся приведенная здесь информация является просто описанием, известным из биологии [14]. Отсюда еще никак не следуют базовые физические принципы функционирования живых систем. Эта задача относится к основным принципам биофизики.
Физические принципы существования живых организмов.
Биофизика, как явствует из ее названия, область физики, призванная объяснять явления биологии с помощью физических принципов. Следовательно, можно ожидать, что пользуясь принципами биофизики, можно значительно облегчить нашу задачу - предсказание биологического действия лазерного излучения.
В действительности положение дел несколько более сложно. Дело в том, что как физическая дисциплина, биофизика не только далека от завершения, но даже сами основные принципы этой науки пока остаются спорными. В результате биофизика не является единой наукой, исходящей из общих законов, а скорее представляет собой набор моделей, более или менее правильно описывающих частные явления.
Совершенно очевидно, что все биологические объекты всегда и полностью подчиняются общим законам природы, в том числе законам физики. Тем самым, физика полностью применима для описания явлений в биологии. Другой вопрос, является ли это описание достаточным, или должны быть введены какие-то дополнительные новые базовые положения биофизики. В 70 - 80 годы доминировал взгляд, что законов физики достаточно для объяснения биологических явлений.
В настоящее время большинство исследователей склоняются к противоположной точке зрения. В качестве примера можно привести позицию патриарха отечественной биофизики Л.А. Блюменфельда, с которым автору посчастливилось быть лично знакомым: если в статьях 70-х годов Блюменфельд отстаивал сведение биологических явлений к чисто физическим процессам, то к концу жизни он явно пересмотрел свои взгляды [15].Другая сложность, лежащая в основании биофизики, связана с различием в методах анализа в биологии и в физике. Очевидно, биофизика должна опираться на данные, полученные в биологии.
Полученные в опытах данные, естественно, должны обобщаться для выяснения более общих закономерностей. Это одинаково в физике и биологии. Но метод оказывается разным: так, в биологии принято так называемое финалистическое описание, т.е. объяснение по принципу «это для того, чтобы..»; в физике, напротив, описание должно быть каузальным (т.е. причинно-следственным) - «это потому, что..». К счастью, это противоречие, как показал М.В. Волькенштейн, скорее кажущееся: так, в физике, любой закон, формулирующийся с помощью какого-либо вариационного принципа, также может быть сведен к биологической форме «для того, чтобы..». (Например, по принципу Ферма, свет «выбирает траекторию на границе двух сред так, чтобы время пути было минимальным.. » и т.п.). Заметим, что в основе всех пяти основных физических теорий лежат некие вариационные принципы. Видимо, так должно быть и в биофизике. Другое дело, что соответствующие биологии вариационные принципы пока еще не полностью определены. Однако изложим то, что уже ясно.
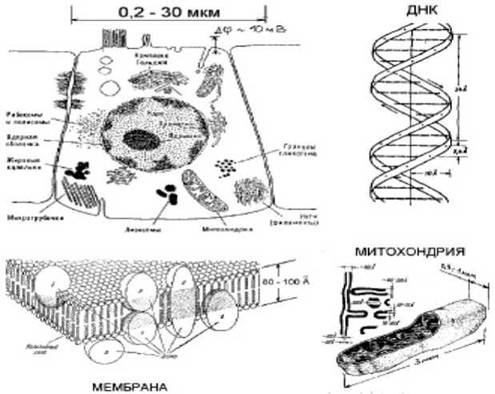
Рис. 7.1. Схема строения клетки, мембраны и митохондрий 135
С точки зрения физики биологические объекты представляют собой самоподдерживающиеся системы взаимосвязанных химических реакций, открытые для энерго- и массобмена с внешней средой.
Долгое время было неизвестно, возможна ли в принципе самоорганизация (самопроизвольное увеличение упорядоченности) в каких- то системах химических реакций.
Дело в том, что организованное состояние материи означает относительный минимум энтропии, тогда как по второму началу термодинамики в любых процессах энтропия должна только расти. В большинстве известных химических реакций именно так и происходит: система стремится к минимуму свободной энергии, т.е. к максимуму энтропии. Поэтому явилось открытием обнаружение т.н. реакции химических часов, или реакции Белоусова - Жаботинского, в которой наблюдалась временная самоорганизация (автоколебания) и, позже, пространственная самоорганизация (образование пространственных структур). По сути, эта реакция моделирует физические принципы существования живых организмов. Рассмотрим схему этой реакции [16].Реакция химических часов (Белоусова - Жаботинского). Реакция химических часов исторически была вначале открыта чисто экспериментально при изучении растворов солей церия. Лишь значительно позже появилась интерпретация, основанная на принципах теории нелинейных колебаний.
Упрощенно и схематически система реакций может быть представлена следующими четырьмя уравнениями:
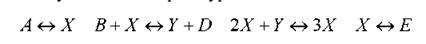
Здесь важно то, что для осуществления реакции концентрации начальных и конечных продуктов должны поддерживаться постоянными в пространстве и времени. Точнее, А, В, D, E поддерживаются постоянными во времени и пространстве. Поскольку эти вещества могут поглощаться (производиться) в указанных реакциях, это условие означает, что мы имеем дело с открытой системой, которой всегда надо поставлять или удалять соответствующие реагенты; т.е. системой, находящейся в энергомассообмене с окружающей средой. Это вполне аналогично обязательно открытому характеру процессов в любом живом организме.
Другой важной чертой является наличие положительной обратной связи в третьей реакции: производство вещества Х пропорционально его же концентрации. В химии такие обратные связи называются автокаталитическими реакциями. Автокаталитическая стадия, включающая тримолекулярную реакцию, вводит в уравнения кинетики нелинейность, которая, как мы видели, является необходимым условием появления неустойчивостей.
Положим для простоты все кинетические константы равными
единице. Отклонение состояния системы от равновесия задано тогда отношениями концентраций А/Е и В/D. Равновесие соответствует равенству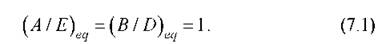
Изучение системы сильно упрощается в предельном случае 
Физически это означает, что продукты D и E удаляются, как только они возникают. Степень отклонения системы от равновесия (количественно характеризующаяся в химии понятием сродства) стремится при этом к бесконечности, т.е. система действует на «бесконечном расстоянии» от равновесия. Кроме того, примем другое упрощение, состоящее в пренебрежении обратными реакциями в оставшихся двух стадиях. Схема, таким образом, становятся полностью необратимой. В этом случае уравнения химической кинетики становятся очень простыми:
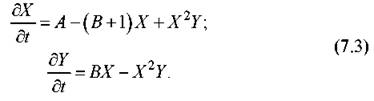
Равновесное решение, т.е. особая точка системы, дается просто стационарным решением

Проводя исследование на устойчивость состояния равновесия согласно методам, изложенным в предыдущей главе, легко находим, что для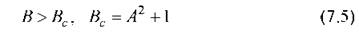 состояние (7.4) неустойчиво. Вслед за неустойчивостью система эволюционирует к незатухающему периодическому режиму, представленному на рис. 7.2.
состояние (7.4) неустойчиво. Вслед за неустойчивостью система эволюционирует к незатухающему периодическому режиму, представленному на рис. 7.2.
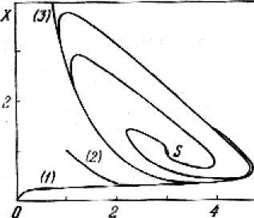
Исследование на устойчивость и анализ фазового портрета системы (7.1) мы оставляем читателю в качестве упражнения. Из рис. 7.2 видно, что этот незатухающий режим достигается всегда, независимо от начальных условий. Его характеристики, например период или амплитуда колебания, являются, следовательно, внутренними свойствами системы и не зависят от начальных условий: напомним, такое поведение известно в нелинейной математике как предельный цикл.
Z
Рис. 7.2. Фазовые траектории в реакции химических часов: формирование предельного цикла
(А = 1, В = 3); S - особая точка; (1), (2), (3) - траектории для разных начальных условий
Тем самым, в системе химических реакций (7.1) возникает самоорганизация во времени - предельный цикл. Главные условия существования этой самоорганизации - наличие автокаталитической стадии (т.е. положительной обратной связи) и открытость системы.
Несколько позже было экспериментально показано, что в системе реакций (7.1) возможна также пространственная самоорганизация. Эта самоорганизация выражалась в формировании сложных двух- и трехмерных структур с упорядоченным пространственным расположением цветов раствора. Для понимания этого явления оказалось достаточно учесть диффузию реагентов в пространстве. Предположим для простоты, что система эволюционирует в единственном пространственном измерении r. Уравнения химической кинетики, соответствующие системе реакции (7.1) с учетом диффузии, тогда имеют вид:
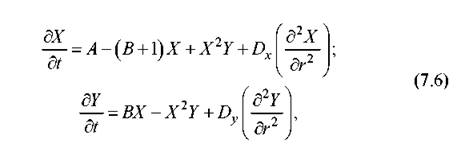
где Dx, Dy - коэффициенты диффузии реагентов X и Y. В этом случае получение аналитических выражений значительно усложняется и характерные пространственные распределения обычно вычисляются численными методами. Физически понятно, что при неоднородном распределении реагентов в пространстве в каждой точке пространства реакция будет находиться в той стадии, которая определяется концентрациями веществ в данном месте. По прошествии времени, даже если везде система реакций уже вышла на предельный цикл, «фаза» этого цикла (конкретная точка на предельной кривой на рис. 7.2) будет различна. В итоге и формируется упорядоченное пространственное распределение вещества. Другой причиной пространственной самоорганизации, как выяснилось, может быть установление новых обратных связей между диффузионными потоками.
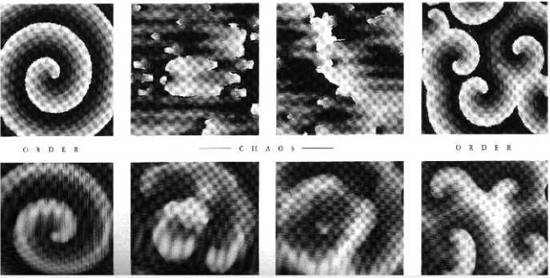
Рис. 7.3. Пространственная самоорганизация в реакции Белоусова - Жаботинского: переходы порядок-хаос-порядок
Примеры пространственных распределений в реакции Белоусова - Жаботинского приведены на рис. 7.3. Как видно из рисунка, при изменении параметров системы, она может последовательно эволюционировать от порядка к хаосу и затем к новому порядку.
Заметим, что все эти состояния являются существенно неравновесными. Неравновесность поддерживается открытостью системы. Подобные свойства в значительной степени характерны и для живых организмов.
Некоторые другие физические принципы теоретической биофизики. Тем не менее ясно, что от реакции Белоусова - Жабо- тинского еще бесконечно далеко до описания живого организма. Даже если считать, что основной принцип найден, то большая сложность биологических объектов все равно не позволит детально проанализировать поведение системы. Иными словами, количество связанных химических процессов в клетке столь велико, что должно характеризоваться нереально большим (макроскопическим) количеством параметров.
Но подобная ситуация уже встречалась в физике. Так, из ньютоновых законов для макроскопических количеств молекул, в статистической физике приходят к ограниченному числу макропараметров - температура, давление, энтропия и т.п. и в дальнейшем получают закономерности, связывающие только эти макропараметры.
Такая попытка перехода к макропараметрам в нелинейных системах химической кинетики, т.е. построение термодинамики открытых неравновесных систем, была осуществлена в работах Пригожина и Николиса еще в 70-е годы XX в. Как известно, из кинетических характеристик можно составить функцию, имеющую физический смысл энтропии, сродство также можно полагать термодинамическим параметром и т.п. Целью этих работ было установление закономерности, выражающей условие существования неравновесных стационарных состояний через такие термодинамические параметры.
Первый принцип, который был получен на этом пути звучит как
условие минимума продукции энтропии. Для систем, находящихся вблизи равновесного состояния, функция производства энтропии в единицу времени («продукция энтропии») в неравновесном стационарном состоянии должна принимать минимальное значение. Для систем, далеких от равновесия вторая вариация энтропии также должна быть отрицательной при достижении стационарного неравновесного состояния. Тем самым, как видим, были сформулированы некоторые недостававшие вариационные принципы биологической физики.
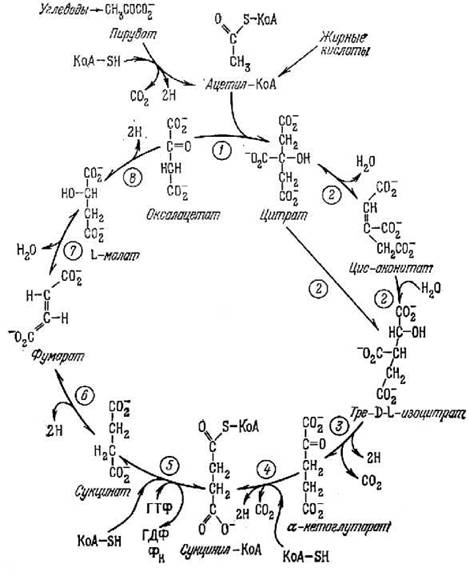
Рис. 7.4. Биохимическая схема цикла Кребса
Эти принципы до сих пор лежат в основе теоретической биофизики. Однако, их использование по-прежнему затруднено сложностью даже элементарных процессов, таких, например, как ферментативный катализ и др. Заметным успехом в использовании этих принципов было построение математической теории эволюции Манфредом Эйгеном в 70е годы XX в. Правда, для этого ему пришлось вводить не только понятие информации, но и довольно умозрительное понятие «ценности информации», которое делает всю теорию чисто феноменологической.
Итак, основные физические принципы функционирования живых систем можно считать более или менее установленными. Для нашей задачи - выяснения механизмов биологического действия лазерного излучения - принципиальное значение имеет получаемое отсюда качественное объяснение пространственной и временной
самоорганизации в системах химических реакций.
Для нас здесь важно то, что подавляющее большинство процессов в живой клетке, как теперь установлено, имеют циклический характер. Так, основные химические процессы в клетках - процессы ферментативного катализа - вообще характеризуются такой величиной, как «число оборотов реакции в единицу времени».
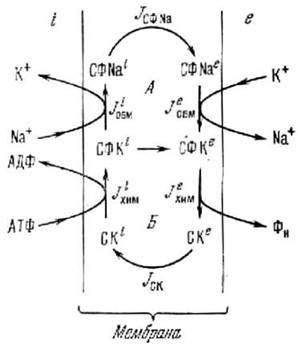
Рис. 7.5. Схема работы мембранного натриевого канала
Для примера приведем еще несколько фактов временной самоорганизации в живых системах. На рис. 7.4 показан так называемый цикл лимонной кислоты (или цикл Кребса). Этот цикл входит в дыхательную цепь высших организмов. Механизм этого цикла был установлен экспериментально. Не следует думать, что временная цикличность характерна только для внутриклеточных реакций. Мембранные процессы, как оказалось, тоже цикличны. Для примера на рис. 7.5 приведен цикл процессов функционирования натриевого канала в мембране клетки [14].
Физические механизмы биологического действия лазерного излучения. Итак, приведенные сведения прямо подсказывают принципиальный физический механизм биологического действия лазерного излучения. Читатель, вероятно, уже заметил прямую аналогию изложенных механизмов с механизмами предыдущей главы. В самом деле, биологические процессы представляют собой циклические химические реакции, вызванные существованием обратных связей и нелинейностью открытой системы. Действие же излучения на химические реакции, как мы видели выше, тоже чаще всего определяется возникновением обратных связей между концентрациями реагентов и параметрами излучения.
Таким образом, базовый механизм биологического действия лазерного излучения можно сформулировать так: лазерное излучение, в принципе, может приводить к образованию дополнительных обратных связей в системах биохимических реакций и, за счет этого, вызывать усиление или подавление протекающих процессов.
Однако, несмотря на принципиальную простоту и ясность этого объяснения, такой механизм оказывается реально подтвержденным лишь в ограниченном числе случаев. Важнейшим, как видели, условием возникновения обратной связи между реакцией и излучением является резонансность поглощения. Посмотрим теперь, какие вещества в клетках могут резонансно поглощать, например, излучение видимого, УФ- и ИК-диапазона.
Проникновение и поглощение излучения. Фосфолипиды являются алифатическими (т.е. непредельными, не имеющими кратных связей) углеводородами. Это означает, что полосы электронного поглощения для них лежат уже на краю ВУФ-диапазона (< 150 нм) и недоступны большинству современных лазеров. То же относится к
воде и почти всем аминокислотам белков. Лишь три аминокислоты
имеют полосы поглощения в ближнем УФ-диапазоне (~ 250 - 310 нм): триптофан (Trp), тирозин (Tyr) и фенилаланин (Pha). Заметим, что эти аминокислоты присутствуют далеко не во всех белках.
Другими акцепторами излучения видимого диапазона могут являться комплексы белков с ионами металлов, например, гемоглобин. Гемоглобин является основным переносчиком кислорода в высших организмах. Спектры поглощения окисленных и восстановленной форм гемоглобина приведены на рис. 7.6. Однако итоговый коэффициент поглощения крови оказывается не слишком высоким (десятки см-1).
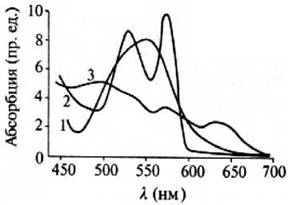
Рис. 7.6. Спектры поглощения дезокси- (1), окси- (2) и ферригемоглобина (3 )
Еще одним поглощающим веществом в красной области видимого диапазона могут являться производные порфирина, которые также присутствуют в достаточных количествах далеко не всегда.
В области ближнего ИК-диапазона (~ 1 мкм) существует область относительной прозрачности (< 1 см-1) большинства биотканей. В области валентных (~ 2 - 5 мкм) и деформационных (~ 5 - 20 мкм) колебаний молекул всегда присутствуют довольно интенсивные (> 50 см-1), но уширенные полосы O-H, C-H, C-N и другие молекулярных колебаний.
Итак, как видим, лишь ограниченное количество молекул может обеспечить резонансное поглощение и формирование описанного механизма обратной связи. Тем не менее, этот механизм все-таки может быть доминирующим. Его, например, всегда можно стимулировать путем разработки соответствующих способов доставки излучения до объекта терапевтического воздействия (например, облучение крови в системе с двумя катетерами и т.п.). Заметим также, что в силу свойств этого механизма (а именно, обратных связей), даже чисто тепловое воздействие лазерного излучения (т.е. при отсутствии микронеравновесности) может приводить к очень оригинальным результатам, вовсе не эквивалентным обычной гипертермии с помощью СВЧ-источников.
Но большинство практических задач лазерной медицины связано с воздействием in vivo (т.е. непосредственно на живую ткань). Здесь возникает другая трудность: если поглощение резонансно, то глубина поглощения света оказывается малой, точнее, чем больше поглощение, тем менее глубоко свет может проникнуть в ткань. Так, например, излучение четвертой гармоники неодимового лазера (266 нм) практически полностью поглощается в эпидермисе (омертвевший верхний слой кожи), и, очевидно, не вызывает биологических эффектов.
Тем самым, мы столкнулись с известным парадоксом: для максимального воздействия излучения требуется высокий коэффициент поглощения, но это приводит к очень малой глубине проникновения излучения и, как следствие, малому воздействию на организм in vivo.
Это явилось одной из причин того, что большинство терапевтических эффектов лазерного излучения наблюдают при воздействии почти нерезонансного излучения, преимущественно в красной части спектра.
Другим препятствием для проникновения лазерного излучения в биологические ткани является сильное рассеяние света. Проникая в биоткань или жидкость, свет рассеивается на клетках, мембранах
и органеллах клеток. Так, в большинстве биотканей коэффициенты
рассеяния в оптическом диапазоне составляют 0,5 - 50 см-1. Это означает, что глубина проникновения даже нерезонансного излучения обычно ограничена несколькими сантиметрами.
Дополнительно заметим, что сильное рассеяние света практически исключает какие-либо механизмы воздействия, основанные на когерентности лазерного излучения. Напомним, что когерентность света - свойство световой волны сохранять фазу волны вдоль пути распространения (временная когерентность) и вдоль фронта волны (пространственная когерентность). Ясно, что уже первый слой клеток (или даже эпидермиса) разрушает когерентность светового поля. Правда, следует заметить, что существуют оригинальные механизмы биологического воздействия лазерного излучения, учитывающие фазу волны (А.Н. Малов [17]); однако, пока они остаются несколько умозрительными.
Изложим теперь некоторые экспериментальные факты, касающиеся воздействия лазерного излучения на компоненты клеток.
Лазерное воздействие на белки. Начиная с появления первых лазеров в 60-х годах двадцатого века проводилось большое количество экспериментов по воздействию лазерного излучения на белки в растворах. Было поставлено множество работ по ультрафиолетовому лазерному воздействию на белки, содержащие резонансно поглощающие аминокислоты - триптофан, тирозин, фенилаланин; по нерезонансному воздействию и по резонансному инфракрасному воздействию с возбуждением ОН, СН и NH валентных или деформационных колебаний. Как показали эксперименты, в подавляющем большинстве случаев наблюдается один процесс - денатурация белковых макромолекул. Это легко понять, если учесть, что в нативной рабочей конформации белок может находиться лишь в довольно узком диапазоне условий (температуре, рН среды и т.п.). Вспомним, что любое возбуждение в конденсированных средах (см. гл. 2 и 3) так или иначе быстро релаксирует в тепло. Денатурация белка как раз и представляет собой переход из рабочей глобулярной конформации (т.е. довольно плотной) в состояние статистического клубка. Примером денатурации может являться изменение состояния яичного белка при нагреве. Таким образом, основное действие лазерного излучения в этих случаях - разрушение рабочей конформации белков.
В середине восьмидесятых годов появилась новая интересная гипотеза о возможности существования колебательных солитонов в макромолекулах белков (так называемые солитоны Давыдова). Напомним, что солитон представляет собой устойчивую уединенную волну, обладающую свойствами, подобными частицам. Физическая причина такой устойчивости солитонов - конкуренция дисперсионного расплывания волнового пакета и сжатия, вызванного нелинейностью среды. Давыдов показал, что если в уравнении для колебательного возбуждения в полимерной цепочке, приближенно описываемом квадратичными потенциалами (см. выше - потенциал Морзе), в порядке следующего приближения учесть кубический член, то получится уравнение, известное в теории солитонов как кубическое уравнение Шредингера. Понятно, что существование таких солитонов означает возможность концентрирования энергии возбуждения в каких-то выбранных областях белка на длительное время. Однако многие эксперименты с использованием антистоксова рамановского рассеяния, когерентного антистоксова рассеяния света (КАРС) и т.п. так и не обнаружили солитонов Давыдова.
Другой интересной особенностью белка является принципиальная возможность длительного существования неравновесного распределения колебательного возбуждения между различными структурными элементами макромолекул: альфа-спиралями, складками и т.п. В экспериментах автора в середине девяностых годов было показано (методом антистоксова рамановского рассеяния), что при резонансном лазерном возбуждении ОН-колебаний в альбуминах распределение возбуждения сильно отличается от СН- возбуждения (такого же по всем термодинамическим параметрам). Таким образом, нельзя исключить существования довольно тонких механизмов воздействия лазерного излучения на конформации белков; однако этот вопрос еще далеко не исследован [18].
Лазерная генетика. Вопреки распространенному убеждению, что ультрафиолетовое излучение всегда влияет на ДНК и на геном, следует заметить, что эксперименты, все-таки, в общем случае не подтверждают этого. Правда, еще с прошлого века хорошо известно бактерицидное («убивающее» бактерии) действие ультрафиолетового излучения. Сегодня почти в каждой клинике производят стерилизацию (так называемое «кварцевание») операционных и процедурных помещений с применением ультрафиолетовых ламп (как правило, дейтериевых или ртутных). Такое бактерицидное действие традиционно связывают с разрушением ДНК микроорганизмов, поскольку именно ДНК обладает достаточно большими сечениями поглощения. Это, разумеется, так, но разрушение обычно достигается только очень большими дозами облучения.
Как показывают эксперименты, сходным образом действует и лазерное УФ-излучение. Так, в опытах Chilbert и др. ([19]) под действием эксимерного лазера наблюдалось разрушение конформации двойной спирали в ДНК и образование циклических полинуклеотидов, причем только при очень больших дозах облучения (> 100 Дж на микрообразец). Образование этих продуктов может говорить о возможной предиссоциации полинуклеотидов (помимо тепловой диссоциации) при таком воздействии. Примечательно,
что этот эффект зависит только от дозы, но не от интенсивности
излучения - это проверялось специально.
В 2005 г. в университете Огайо в США (Kohler и др.[20]) с помощь фемтосекундного лазера удалось точно измерить характерные времена жизни электронно-возбужденных состояний в ДНК. Оказалось, что эти времена меньше одной пикосекунды. Авторы сочли это «естественным механизмом защиты ДНК от ультрафиолетового повреждения». Ясно, что интенсивности света, преодолевающие такую быструю релаксацию in vivo недостижимы.
Одна из немногих работ, в которой действительно обнаруживается модификация генома лазерным излучением была проделана в 2005 г. с помощью микроскопического «лазерного прокалывания» (puncture) мембран отдельных зародышевых клеток пшеницы (Badr и др.) [21]. Это делалось с помощью острой фокусировки излучения УФ эксимерного лазера в оптическом микроскопе. При этом мембраны быстро самовосстанавливались, а ДНК ядра получало дозу облучения. В результате появлялись жизнеспособные клетки с измененным геномом. Изменение генома затем исследовалось отдельно. Результат показан на рис. 7.7.
Думается, что этот эффект может быть связан не столько с воздействием непосредственно на ДНК, сколько, скорее, с модификацией каких-то генетических механизмов, обеспечиваемых другими молекулами.
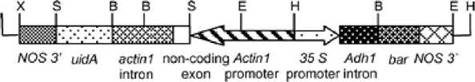
Рис. 7.7. Изменение генома при лазерном «пунктировании» клеток пшеницы (Badr et al, 2005)
Итак, пока каких-то избирательных механизмов модификации генома лазерным излучением, похоже, пока не найдено. Однако
нам уже сейчас ясен основной механизм лазерного действия на
биологические объекты (обратные связи «излучение - продукт») и ясны физические условия его реализации. Может быть со временем такие механизмы будут использоваться в молекулярной генетике; здесь - огромные перспективы. Несмотря на это, пока удручающе мало исследований с применением описанного механизма к конкретным (известным) биохимическим процессам. Зато процветает другая область лазерной медицины: воздействие низкоинтенсивного лазерного излучения.
Терапевтическое действие низкоинтенсивного лазерного излучения. В отечественной медицине считается клинически доказанным лечебное действие низкоинтенсивного лазерного излучения (интенсивности ~ мВт/см2) с длиной волны излучения в красной области (~ 600 - 800 нм). Чаще всего для этих целей используется излучение самых доступных гелий-неоновых лазеров (633 нм) или полупроводниковых лазеров (700 - 900 нм). Терапевтическое действие такого излучения относится к наиболее загадочным вопросам лазерной медицины.
Сразу заметим, что исследования лечебного действия такого излучения и применение его в медицинской практике проводятся только в нескольких странах: в России, и (в меньшей степени) в Израиле, Японии, Китае и Латинской Америке. В США, Канаде и Западной Европе и других странах мира интерес к этим методам практически отсутствует.
Одной из причин этого является тот факт, что эффекты такого воздействия почти неотличимы от эффектов плацебо. В медицине плацебо называется препарат, внешне неотличимый от лекарства (таблеток и т.п.), но не содержащий никаких лекарственных веществ. Прием такого препарата больным, убежденным в том, что это лекарство, как теперь твердо доказано, вызывает заметный лечебный эффект. Причиной этого, по всей вероятности, является психосоматический эффект или самовнушение.
Термин «плацебо» был введен в медицине еще в 1894 г. Однако эффект плацебо начал учитываться только в 30-е годы XX в., после введения предложенного Брэдфордом Хиллом метода рандомизированных испытаний любого лекарственного препарата. Суть метода состоит в том, что действие лекарства на какой-то группе пациентов должно обязательно сравниваться с действием плацебо на контрольной группе. Только различие в результате воздействия на основную группу и контрольную группу пациентов говорит о лекарственных свойствах препарата. Любопытно, что в настоящее время больше половины используемых лекарственных средств так и не прошли рандомизированного исследования. Сейчас (в начале XXI в.) проведены масштабные исследования этого эффекта в США и Европе. Получены интересные результаты: оказалось, что почти все лекарственные препараты, использовавшиеся в XIX в. и не менее половины в XX в. действовали по механизму плацебо. Другой, важный для нас результат состоит в том, что эффект плацебо практически не работает при лечении инфекционных заболеваний, а также травм, ран (2) и т.п.
Конечно, рандомизированные исследования в нашей стране проводились и по отношению к низкоинтенсивной лазерной терапии. Результат оказался разочаровывающим. Обобщение большого материала, основанного на применении плацебо (когда больной проходил как бы весь курс лечения, но лазер не включали) и двойного слепого контроля (когда о том, где плацебо, а где лазер, не знали ни больной, ни лечащий врач), показало во многих случаях отсутствие статистически достоверных улучшений в состоянии больного. В тех же случаях, когда лечебный эффект был, врачи жаловались на очень плохую воспроизводимость результатов, различия в чувствительности к облучению у разных больных, опасность передозировки, когда лечебный эффект облучения сменялся неблагоприятным действием.
Итак, с высокой вероятностью, механизм лечебного действия низкоинтенсивного лазерного излучения может сводиться к эффекту плацебо. На этом можно было бы закрыть эту тему, однако, как оказалось существуют некоторые факты, показывающие, что лечебное действие такого излучения действительно возможно. Во- первых, большое число успешных применений излучения относилось к заживлению послеоперационных ран, когда, как упоминалось выше, обычный эффект плацебо обычно не работает. Во- вторых, биохимический анализ клеток, подвергавшихся облучению in vitro, объективно показывает некоторое изменение химического состава. Есть ряд других, еще более косвенных подтверждений возможности лечебного эффекта низкоинтенсивного лазерного излучения, не связанного с эффектом плацебо.
В связи с этим, а также с высокой вероятностью встречи в практической работе нашего специалиста с такими методами лечения, приведем объективные факты и основные гипотезы низкоинтенсивной лазерной терапии.
Прежде всего, приведем список примеров успешного применения такой лазерной терапии: облучение крови и суставов при ревматоидном артрите, лечение мужского бесплодия, глазных болезней, включая глаукому, дисфункции мочевого пузыря, ишемической болезни сердца, кожных болезней и болезней крови, осложнений при хирургических травмах, пиелонефрита, плацентарной недостаточности, туберкулеза легких, подготовка больных к хирургическим вмешательствам, лечение опухолей и др.
По мнению врачей, главной проблемой в области лазерной терапии является отсутствие научного объяснения наблюдаемых положительных (и отрицательных) эффектов лазерного облучения. По формулировке Ю.А. Владимирова из РГМУ [22], «не понимая, что происходит, нельзя правильно лечить».
Объективно установлены следующие факты [22].
1. Излучение с длиной волны 400 - 500 и около 600 нм вызывало увеличение скорости деления клеток некоторых микроорганизмов, а также увеличение белкового синтеза. При этом наблюдалась выраженная экстремальная зависимость стимулируюшего действия лазерного облучения от дозы; интервал интенсивностей, в пределах которых наблюдался положительный эффект, составлял полтора- два порядка величины.
2. При облучении изолированных митохондрий печени светом гелий-неонового лазера в них увеличивалось содержание АТФ, электрический потенциал на внутренних мембранах и рН в матриксе, а также наблюдаются небольшие изменения формы матрикса.
3. Облучение клеток-фибробластов человека светом гелийнеонового лазера существенно увеличивало скорость роста популяции клеток в экспоненциальной фазе (как говорят, пролиферацию клеток) и (в случае эмбриональных фибробластов) адгезию клеток на подложке.
4. Лазерное облучение клеток, выделенных из эпителия сетчатки, приводило к увеличению митотического индекса (митотический индекс - показатель митотической активности ткани или культуры ткани, представляющий собой число делящихся клеток из 1000 изученных на гистологическом препарате.).
5. Под действием облучения гелий-неонового лазера увеличивалось содержание внутриклеточного кальция в лейкоцитах, окрашивание нуклеиновых кислот в клетках флуоресцирующим красителем (акридиновым оранжевым) и изменение морфологической структуры хроматина. По всем параметрам действие лазерного излучения в оптимальной дозе было сходным с действием вещества - фитогемагглютинина.
6. При облучении крови непосредственно в кровеносных сосудах наблюдалось расширение сосуда выше и ниже места облучения; эффект исчезал при замене крови перфузируемым физиологическим раствором. Этот эффект (вазодилатация) приводит к улучшению кровоснабжения органов.
Таким образом, все наблюдаемые в клинике эффекты (противовоспалительное действие лазерного облучения, ускорение регенерации поврежденных тканей и улучшение кровоснабжения органов) могут быть связаны с тремя эффектами, полученными в эксперименте:
1) ростом активности фагоцитов;
2) усилением пролиферации клеток;
3) улучшением циркуляции крови по сосудистому руслу вследствие расслабления стенок кровеносных сосудов (вазодилатации).
Главная проблема, не позволяющая пока детально объяснить эти наблюдения - отсутствие какого-либо вещества, заведомо участвующего в этих процессах и эффективно поглощающего излучение гелий-неонового лазера. Несмотря на неясность с акцептором излучения, далее предполагаются чисто химические механизмы (что естественно, поскольку эти исследования находились в руках медиков). Фотохимические механизмы, которые могут обеспечить протекание описанных процессов, в настоящее время более или менее ясны:
1) фотоокисление липидов в клеточных мембранах;
2) фотореактивацию фермента супероксиддисмутазы (СОД);
3) фотолиз комплексов окиси азота (NO).
Как уже отмечалось, чистые фосфолипиды не поглощают свет в видимой и ближней инфракрасной областях спектра, характерных для излучения использованных лазеров. Однако, если к фосфолипидам добавить поглощающий свет сенсибилизатор, например гематопорфирин или фталоцианин, то под действием света лазера, в частности, гелий-неонового, происходит окисление липидов (фотодинамический эффект), сопровождающееся образованием перекисей (фотопероксидация). Фотопероксидация липидов наблюдается также и при лазерном облучении суспензии клеток или митохондрий, всегда, если добавить сенсибилизатор, а иногда, если даже его не добавлять. По-видимому, в некоторых условиях, в частности при определенных заболеваниях, в мембранах накапливаются природные сенсибилизаторы, возможно тот же гематопорфирин или его производные.
Итак, если при каких-то, пока не совсем ясных условиях, возникает накопление природного фотосенсибилизатора, то дальнейшие биохимические эффекты, в общем, на сегодня понятны. Добавим, что изучение этих механизмов позволило объяснить наиболее характерную черту низкоинтенсивной лазерной терапии: кривые зависимости эффект-доза, имеющие максимум. Характерный вид кривых показан на рис. 7.7. Эффективность воздействия в этих экспериментах контролировалась по величине хемилюминесценции лейкоцитов крови.
Как видим, результаты, в общем, согласуются с выработанными представлениями о существовании естественного сенсибилизатора. Проблема в том, что эти объяснения далеко не единственны. Приведем для ознакомления другие модели, столь же успешно объясняющие лечебное действие низкоинтенсивного лазерного излучения.
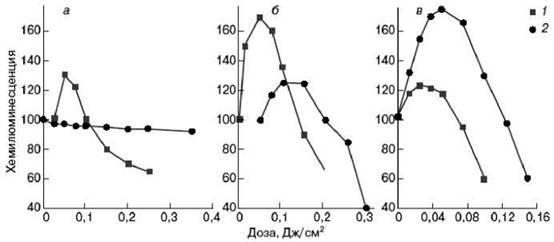
Рис. 7.7. Лазерная стимуляция (прайминг) лейкоцитов, изолированных из крови
больных различными легочными заболеваниями, вызванный лазерным облучением: а - хронический бронхит; б - хроническое воспаление легких; в - тяжелая полисегментная пневмония в острой фазе; 1 - в присутствии искусственного сенсибилизатора; 2 - без сенсибилизатора
Предполагается, что акцепторами лазерного излучения могут быть:
1) порфирин и его производные [23, 24, 25];
2) молекулы ферментов-антиоксидантов: супероксид-дисмутаза (СОД), каталаза, церулоплазмин [26, 27];
3) компоненты дыхательной цепи митохондрий: флавопротеины и цитохромы [24, 28];
4) молекулярный кислород [30, 31].
Эти вещества, конечно, могут быть акцепторами излучения гелий-неонового лазера, однако сечения поглощения при этом для них (за исключением порфиринов) довольно малы. Суммарное количество поглощенных квантов будет столь невелико, что концентрация продуктов соответствующей фотохимической реакции должна быть пренебрежимо мала (по сравнению с обычной химической терапией). Другим контраргументом является тот факт, что сообщалось и о терапевтическом действии низкоинтенсивного лазерного излучения других длин волн: в зеленой части спектра (514 нм) и инфракрасной (950 - 1000 нм). В этом случае эффект уже не может связываться с поглощением порфиринами и т.п.
Также предлагается большое количество механизмов, приводящих к терапевтическому эффекту:
1 - реактивация металлосодержащих ферментов-антиоксидантов [31, 32, 26, 27];
2 - гипотеза о взаимодействии низкоинтенсивного лазерного излучения с компонентами цепи транспорта электронов в митохондриях [21, 28];
3 - неспецифическое влияние на биополимеры [26];
4 - фотовозбужденное образование синглетного кислорода [29,
30];
5 - неспецифическое влияние на структуру воды [30].
Мы не будем подробно рассматривать все эти механизмы, отметим только общие черты: во-первых, все эти механизмы удовлетворительно объясняют наблюдаемые эффекты; во-вторых, против всех этих механизмов существуют очень серьезные контраргументы; в-третьих, существуют факты, подтверждающие тот или иной механизм, но носящие обычно косвенный характер.
В заключение приведем еще одно очень простое соображение. Мы видели, что когерентность лазерного излучения не может быть существенной для лечебного эффекта (ее просто нет при распространении света в биотканях). Тогда, единственное, чем может
отличаться лазерное воздействие от простого солнечного света - это спектральная яркость излучения. Но мы видим, что эффекты
вовсе не специфичны по длинам волн, т.е. почти не зависят от спектра света. Заметим, что в яркий солнечный день интенсивность света (до 50 мВт/см2) может быть даже выше интенсивности, скажем, гелий-неонового лазера. Однако, почему-то никто не считает, скажем, простое пребывание в освещенной комнате в течение нескольких минут медицинской процедурой, эффективно излечивающей массу серьезных заболеваний.
Практические применения лазерного излучения в медицине.
Помимо рассмотренного терапевтического действия низкоинтенсивного лазерного излучения, существует ряд более простых и общепризнанных применений лазеров в медицине.
Механическое действие лазерного излучения, «лазерный скальпель». При достаточно высоких интенсивностях излучения, в доплазменном режиме или даже выше порога плазмообразования, нагрев поверхности может достигать столь высоких величин, что вызывает фазовые переходы, испарение или последующую ионизацию вещества в лазерном поле (с образованием факела плазмы). Это приводит к абляции (т.е. «выносу») большого количества вещества с поверхности. Метод удаления вещества с помощью интенсивного лазерного излучения используется в хирургии в качестве скальпеля. Первичный физический процесс здесь, по-видимому, все-таки чисто тепловой. Это подтверждается тем, что наиболее эффективно для этих целей непрерывное или импульснопериодическое излучение, которые приводят к значительному нагреву. Лазерная специфика здесь связана лишь с двумя факторами: монохроматичностью одномодового излучения позволяющей получить очень малые размеры пятна фокусировки излучения и импульсно-периодическим режимом при наносекундных импульсах, что позволяет создать режим сильного нагрева поверхности, при котором средний нагрев ткани не слишком велик, но на поверхности достаточен для протекания абляции. Это выгодно отличает лазерный скальпель от, например, радиочастотного ВЧ-скальпеля.
В медицинских публикациях сообщается о значительно лучшем заживлении послеоперационных ран при использовании лазерного скальпеля вместо обычного. Причины такого эффекта пока не очень ясны. Возможно, при использовании излучения просто меньше травмируются окружающие ткани. Чисто плазмохимическое действие лазерного излучения пока мало изучалось.
Добавим, что эти эффекты (т.е. разница в действии лазерного и обычного скальпеля) обычно не очень велики и часто нестабильны. Если учесть, что лазерная техника является довольно дорогостоящей, то понятно, что вряд ли следует ожидать бурного развития таких методов. Заметим, что, пользуясь методами, изложенными в предыдущих главах, читатель вполне сможет моделировать такое действие излучения.
Лазерная фототерапия и фотодинамическая терапия. В фотодинамической терапии обычно используется какая-либо заранее известная фотохимическая реакция (например, одноквантовая фотодиссоциация связей или химическая реакция молекул в возбужденном состоянии) в определенном веществе, которое вводится в организм извне. Подобные методы уже имеют успех, например, в онкологии. Чаще всего в организм вводятся молекулы производных порфирина, накапливающиеся, как показано, в определенных клетках, которые следует разрушить (например, в метастазирующих тканях). Из этого механизма (определенно одноквантового) ясно, что эффект пропорционален только дозе облучения, но не интенсивности. (Выход одноквантового эффект, конечно, растет с увеличением интенсивности, но только за счет того, что при большей интенсивности за то же время облучения набирается большая доза. Иными словами, при фиксированной дозе эффект не зависит от интенсивности излучения.) Это означает, что использование лазерного излучения в фотодинамической терапии могло бы дать какое-то количественное, но
не качественное преимущество. Однако, как оказалось, этого тоже
не происходит - при помощи ламп дозу можно набрать даже быстрее. Поскольку для фотодинамической терапии не требуются высокие интенсивности излучения, то, возможно, монохроматичность лазерного излучения дает какой-то выигрыш в эффекте? Нет, поскольку сложные молекулы и так обладают широкими полосами поглощения. Как следствие, использование значительно более дешевых ламповых источников света с широким спектром позволяет даже легче набрать необходимую терапевтическую дозу облучения.
Таким образом, использование лазеров в фотодинамической терапии пока также не очень перспективно.
Резюмируем сказанное. Итак, если оставить в стороне простейшие применения лазеров, то лазерное воздействие на биологические объекты по механизму, аналогичному лазерному управлению химическими реакциями (образование новых обратных связей и т.п.) имеет огромные перспективы применений в медицине. Однако здесь необходимы серьезные фундаментальные исследования. К сожалению, в нашей стране они пока почти полностью подавляются гиперактивностью в области низкоинтенсивной лазерной терапии. Проблема здесь, конечно, не в том, что широко проводятся именно такие исследования. Просто с течением времени, очевидно,
возникнет неизбежное разочарование в возможностях низкоинтенсивной терапии (как говорят, сменится мода) и это - столь же неизбежно! - вызовет тотальное недоверие вообще ко всем методам лазерной терапии. Естественно, этого хотелось бы избежать.