ГЛАВА IX ОБЩЕБИОСФЕРНЫЕ ЭКОЛОГИЧЕСКИЕ ФУНКЦИИ ПОЧВЫ
В данной группе глобальных функций почвы мы выделяем в числе первых функцию среды обитания для организмов суши, без которой трудно себе представить полноценную биосферу. Огромное значение почвы как среды обитания для растений и животных Земли проявляется прежде всего в том, что именно с ней связано существование большинства видов живых организмов и образование основной массы живого вещества планеты (табл.
46).Еще сравнительно недавно полагали, что основная биомасса Земли сосредоточена в Мировом океане. Такое суждение основывалось прежде всего на доминировании водной поверхности над сушей. Однако более точные подсчеты показали, что
масса живого вещества континентов многократно превышает биомассу океана. Объясняется это в первую очередь большой плотностью жизни в наземных биогеоценозах, в том числе в почвенной оболочке земли. Так, подсчеты распределения зоомассы на суше показывают, что большая ее часть приурочена к почвенному ярусу в связи с исключительной его насыщенностью жизнью (Дювиньо, Танг, 1968).
Таблица 46
Масса живого вещества Земли (по данным Н. И. Базилевич, Л. Е. Родина, Н. Н. Розова)
| Показатель | Фитомасса | Консументы, редуценты | Итого |
| Континенты | 2,400-1012 | 0,020-1012 | 2,4200-1012 |
| Мировой океан | 0,0002- 10»а | 0,003-1012 | 0,0032-1012 |
| Всего живого вещества | 2,4232-1012 |
О важности и масштабах проявления почвенной функции аккумулятора и источника вещества и энергии для организмов суши можно судить по круговороту и накоплению органического вещества в биосфере (табл.
47—49; схемы III, IV) (Ковда, 1975; Гришина, 1976; Горшков, и др., 1980; Будыко, 1984; и др.).Подсчеты М. М. Кононовой (1974) показывают, что в почвенной оболочке Земли сосредоточено около 2500 млрд, т гумуса. Распределены запасы гумуса по зонам неравномерно. Около половины их приурочено к ферраллитным и черным тропическим почвам (табл. 48). В черноземах, несмотря на их ограниченные площади, сосредоточено более 200 млрд, т гуму-
Таблица 47
Затраты энергии на почвообразование и энергия, аккумулируемая в растительном веществе, ккал/см2/год (Волобуев, 1960)
| Природные зоны | Общая затрата энергии на почвообразование (включая эвапотранспи- рацию) | Энергия, ежегодно аккумулируемая в растительном веществе |
| Тундра и пустыни Черноземные степи Влажные субтропики Влажные тропики | 1 000— 5 000 15 000—30 000 40000—50 000 50 000—70 000 | 5— 60 150— 360 600—1 500 1 500—3 500 |
са; в подзолистых почвах — 183 млрд. т. Ежегодное образование гумусовых веществ в пересчете на углерод составляет 1— 2 млрд. т. Период формирования запасов гумуса составляет 800—1500 лет (табл. 49).
Таблица 48
Запасы гумуса почв мира (Кононова, 1974)
| Почва | Запас в слое 0—1 м, т/га | Площадь ПОЧВ, Л-10’1 млрд. • га | Общие запасы в слое 0—1 м, млрд, т |
| Тундровые и полярных пустынь .... | 73 | 8,1 | 59 |
| Подзолистые................................................. | 99 | 18,7 | 185 |
| Серые лесные............................................... | 215 | 4,5 | 97 |
| Бурые лесные............................................... | 263 | 7,4 | 195 |
| Черноземы.................................................... | 500 | 4,1 | 205 |
| Каштановые................................................. | 160 | 3,6 | 58 |
| Сероземы» серо-бурые, такыры................. | 40 | 31,0 | 124 |
| Красноземы, желтоземы.............................. | 282 | 6,2 | 175 |
| Коричневые.................................................. | 253 | 7,3 | 188 |
| Ферралитные, черные тропические .... | 300 | 42,5 | 1275 |
| Суммарные и средние показатели .... | 192 | 133,4 | 2561 |
Таблица 49
Приблизительные расчеты новообразования гумуса (Кононова, Александрова, 1974)
| Запас в почвах мира, млрд, т | Ежегодное поступление опада, млрд, т | Ежегодное новообразование гумуса, млрд, т С | Период формирования запасов гумуса, годы |
| Гумус 2500 Сгумус 1500 | Опад 115—117 Сопад —70 | 1—2 | 800—1500 |
В настоящее время в связи с широким освоением почвенного покрова и усилением эрозии почв происходит сокращение мировых запасов гумуса. В год их уменьшается на 1,2— 1,4 млрд, т, а за последние 100 лет потеряно около 400 млрд, т гумуса.
Отмечается также снижение темпов новообразования гумуса из-за уменьшения поступления опада более чем на 40% в связи с сельскохозяйственной деятельностью. В доисторическое время, по данным С. П. Горшкова и др. (1980), новообразование гумуса в биосфере составляло 1,8—3,6 млрд, т в год. Таким образом, наряду с проблемой рационального использования различных минеральных ресурсов перед человечеством возникла не менее острая проблема разумного использования и охраны гумусосферы Земли.В рассматриваемую группу входит также почвенная функция связующего звена биологического и геологического круговоротов. Эти два типа круговоротов во многом принципиально' не сходны (Вильямс, 1949). Одно из главных их различий за-
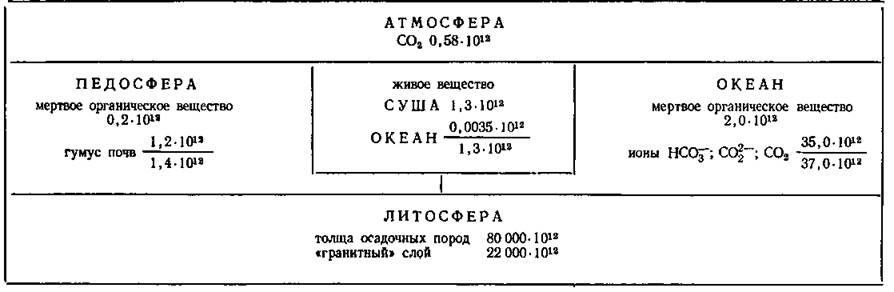
Распределение масс углерода а географической оболочке (цифры—масса элемента, т) (Добровольский, 1984)
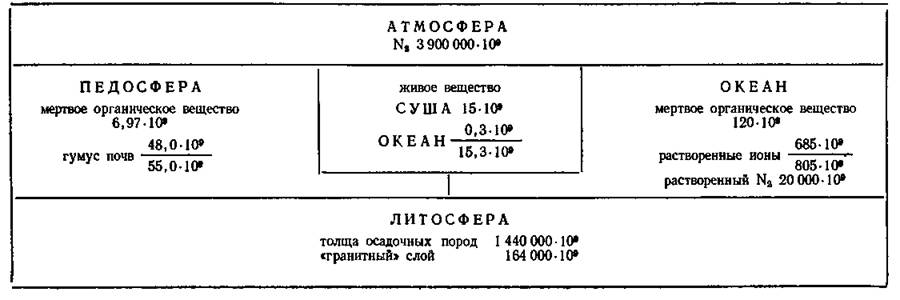
Распределение масс азота в географической оболочке (цифры — масса элемента, т) (Добровольский, 1984)
ключается .прежде всего в темпах и сроках завершения полнено цикла. Геологический круговорот протекает несопоставимо более медленно, чем биологический. Поэтому за относительно короткие сроки, измеряемые годами, десятилетиями, столетиями, на отдельных отрезках геологического круговорота доминирует одно направление потока вещества — снос на плакорах и накопление в акваториях. Биологический же круговорот за те же сроки может обеспечивать полный цикл (от создания биомассы до ее разрушения).
Другое важное различие данных круговоротов на подавляющей части суши заключается в противоречивости их взаимодействия. Биологический круговорот в отличие от континентального отрезка геологического в целом направлен на аккумуляцию и удержание элементов на водоразделах, испытывающих постоянную денудацию. Следует при этом подчеркнуть, что общая накопительная направленность биологического круговорота была бы невозможна без почвы, которая оказывается мощным аккумулятивным и сорбционным барьером на пути мобильных соединений, образующихся после разложения органического опада.
Особую роль здесь играют гумус, органо-минеральные комплексы и вторичные минералы почвы, образование которых приводит не только к связыванию элементов, мобилизованных биологическим круговоротом, но и к их обменному поглощению, являющемуся эффективным механизмом удержания вещества от его выведения из биогеоценозов континентальным геохимическим потоком (Глазовская, 1964; Глазовский, 1976; и др.).Наглядным примером в-ажности почвы в реализации аккумулятивной деятельности биологического круговорота могут служить материалы по биохимическим циклам элементов в различных природных зонах. Исследование Т. И. Евдокимовой, Т. Л. Быстрицкой, В. Д. Васильевской, Л. А. Гришиной, Е. М. Самойловой (1976) показали, что в случае нарушения почвенного покрова происходит принципиальное изменение в соотношении рассматриваемых круговоротов в сторону ослабления биологического и усиления геологического. Так, после распашки лесостепной зоны европейской части СССР геологический круговорот возрос во много раз. Вследствие эрозии почв ежегодно с поверхности водоразделов сносится около 200 млн. т плодородной земли, из которых около 20 млн. т полностью теряется, так как поступает в реки. В доисторический период в биологический круговорот вовлекалось биофильных элементов примерно в 30 раз больше, чем поступало их в геологический круговорот. В современный период количество биофилов, мигрирующих в биологическом круговороте, всего лишь выше их выноса в геологический круговорот.
В степной зоне Русской равнины в доисторическое время вовлечение биофилов в биологический круговорот в 50 раз превышало их поступление в геологический. В настоящее время поступление N, Р, Са, S, Mg в биологический круговорот лишь
зв 2—10 раз выше их выноса поверхностным стоком. Для этих элементов существует угроза быстрой потери их ландшафтами плакоров. Предотвратить данный процесс, компенсируя его только внесением минеральных удобрений, трудно. Существует другой, более действенный, путь — сокращение ветви геологического круговорота за счет полного снятия или значительного уменьшения твердого и жидкого стока, вызываемого водной эрозией.
Для степных пространств, так же как и для лесостепи, это первоочередная задача (Евдокимова и др., 1976).Приведенные примеры показывают, что почвенный покров — важный регулятор взаимодействия биологического и геологического круговоротов и, следовательно, при нарушении почвенной оболочки Земли неизбежно возникают глубокие изменения в сложившихся геохимических потоках биосферы. Таким образом, ненарушенный покров оказывается также защитным барьером и условием нормального функционирования биосферы, в целом. В этом заключается его еще одна весьма важная глобальная экологическая функция.
Одной из наиболее важных является функция почвы как фактора биологической эволюции. До сих пор всесторонняя оценка ее значимости не реализована в полной мере, в том числе среди почвоведов, и особенно специалистов-практиков, которые нередко смотрят на почву лишь как на объект хозяйственного использования. Среди биологов до фундаментальных исследований М. С. Гилярова эволюционному значению почв также не уделялось должного внимания, хотя именно анализ взаимодействия наземных растений и животных с почвенной средой помогает вскрыть и лучше понять многие особенности их развития.
Специфичность почвы, отмечает М. С. Гиляров, как совершенно особого естественно-исторического тела, позволившая В. В. Докучаеву выделить ее в качестве «четвертого природного тела», недооценивается или чаще вовсе не принимается во внимание биологами. Анализ условий обитания животных в почве дает возможность раскрыть исключительное значение особенностей этих условий в эволюции животного мира, в процессе освоения суши исходными организмами. К этому следует добавить, что оценка почвы как основного фактора эволюции органического мира привела некоторых ученых к мысли, что средой зарождения жизни была почва.
К настоящему времени накоплены достаточно обширные сведения, свидетельствующие о выдающемся значении почвы в биологической эволюции биосферы. Поскольку они систематизированы главным образом по животному миру, то роль почвы как ^фактора биоэволюции мы рассмотрим на эколого-зоологическом материале, проанализированном в работах М. С. Гилярова и его школы.
Оценивая роль почвы в эволюционном процессе, М. С. Гиляров (1949) обращает в первую очередь внимание на то, что
почвенная оболочка по своим главнейшим экологическим особенностям может рассматриваться как среда промежуточная (между водной и воздушной), через которую возможен постепенный переход от водного образа жизни к наземному без резкого изменения организации живого. Такая особенность почвы определяется прежде всего водно-воздушными ее свойствами,, по которым она является как бы промежуточной между водой и атмосферой.
Воздух в почве, как правило, насыщен парами воды. Даже в пустыне в жаркий сухой период года в песке на глубине нескольких сантиметров воздух в почве близок к насыщению парами влаги (>90%). Благодаря водонасыщенности воздушной фазы для почвы разрешим Известный физиологический парадокс «дыхание всегда происходит в воде», который связан с тем, что газообмен между поверхностью дыхательных органов, и средою обычно происходит через жидкость.
Отсутствие угрозы гибели от высыхания в почве создает предпосылки для перехода к обитанию в ней водных форм животных с развитой способностью к кожному дыханию, широко распространенной у многих обитателей водоемов. При переходе же к жизни в воздушной среде кожное дыхание становится невозможным. У насекомых препятствием этому служит образование защитного хитинового покрова, в связи с чем функция дыхания должна осуществляться отдельными участками — трахеями, образовавшимися в результате вдавливания дыхательных поверхностей внутрь.
Другой особенностью почвенной среды, способствовавшей переходу водных организмов к обитанию в почве, явилось обилие здесь органического вещества, что, по-видимому, оказалось основной предпосылкой для поселения в ней многих обитателей бентоса.
В результате освоения почвы в ней стали жить представители разных систематических групп животных. Среди беспозвоночных в почве распространены организмы разной степени эволюционной продвинутости.
Широко представлены в почве Protozoa. Причем, в отношении простейших важно отметить, что среди них нет настоящих обитателей воздушной среды. Простейшие либо типичные обитатели водоемов, хотя бы таких, как капли воды в трещинах деревьев, либо почвенные обитатели, находящиеся или в подвижных почвенных растворах, или в состоянии адсорбции на почвенных частицах в пленках воды (Гиляров, 1949). Рассматривая пути перехода от водной среды к почвенной таких представителей простейших, как раковинные амебы (корненожки), исследователи отмечают (Карганова, 1977; и др.), что особенно благоприятные условия постепенного освоения почвы складываются на участках суши, прилегающих к водоемам (озерам и особенно болотам), где всегда имеются частицы ила и органические остатки и где моховые кочки неравномерно и непо-
«6
•стоянно увлажнены. Из 1100 видов корненожек большинство представлено обитателями заросших растительностью пресноводных водоемов и болот, более 11% встречается в почве.
Важным этапом в переходе к наземному (в широком значении слова) обитанию оказалась почва и для различных групп червей. Так, хорошо известна широкая освоенность почвы олигохетами, где выделяются земляные (дождевые) черви, без которых нельзя представить животный мир большинства почв. Среди олигохет, обитающих в водной среде, где преобладают пресноводные формы, также отмечается переход к жизни в почве. В качестве примера можно указать на Enchytracidae, численность которых на 1 м2 выражается нередко тысячами и десятками тысяч.
Отмечен также случай приспособления типично водной олигохеты Tubifex к обитанию в почве. Было установлено, что после осушки одного из озер в почве продолжали жить и размножаться трубочники, в организации которых произошли определенные изменения. Этот случай приспособления к жизни в почве типичного обитателя бентоса убедительно подтверждает возможность перехода водных беспозвоночных к наземному образу жизни через почву.
Широко распространены в почвах и круглые черви — нематоды, которые живут повсюду, в том числе и в почвах сухих зон. Первоначальной же их средой является море.
Среди многощетинковых червей — полихет (Polychaeta) — основная масса — обитатели моря. Однако и здесь были найдены полихеты, переходящие к наземному образу жизни, которые дышат воздухом и обитают в почвах мангровых зарослей.
Подавляющее большинство свободноживущих плоских червей — турбеллярий — являются водными обитателями. Но и ■среди них встречаются формы, переходящие к наземному образу жизни. Причем для неводных турбеллярий характерно обитание именно в почве, а также подстилке. При этом многие ■формы встречаются во влажной почве чаще, чем в воде (Гиляров, 1949). Имеются обитатели почвы и среди немертин, хотя •большинство их — морские формы.
Переход от водного к наземному образу жизни через почву ■отмечен также среди ракообразных, характерной средой которых является море. В качестве примера можно привести наземных мокриц, которые распространены широко и встречаются даже в пустынях (Димо, 1945). Днем, когда угроза гибели от высыхания особенно велика, пустынные мокрицы скрываются в норах.
В процессе освоения суши почва выполняла важную роль переходной среды не только по отношению к беспозвоночным, по и многим позвоночным животным. М. С. Гиляров подтверждает данное положение рядом примеров. Так, известны случаи зарывания в почву некоторых рыб. Например, вьюн проводит сухой летний период, зарывшись в грунт пересыхающего водоема. Лабиринтова рыба Anabas scaundes, имеющая приспособленные к длительному пребыванию вне воды жаберные полости, может переползать в мокрой траве на большие расстояния иэ водоема в водоем. При неблагоприятных условиях анабас закапывается в землю, где длительное время сохраняет жизнеспособность. Частая встречаемость при раскопках послужила причиной того, что эта рыба получила название «земляной рыбы».
В почве также проводят сухой период и двудышащие, зарывшись в высыхающий ил или в землю. Из земноводных после нереста зарываются в землю и зимуют в ней тритоны; проводят сухой период года в земле и жабы.
У рептилий эмбриональный период связан с почвой — в ней осуществляется развитие их яиц.
Роль почвы как переходной среды выявляется и при анализе экологии тех наземных животных, которые в процессе эволюции в наибольшей степени освободились от связей с почвой. Например, даже для такой ксерофильной группы насекомых, какими являются саранчевые, почва продолжает оставаться определенное время средой их обитания — в ней проходит развитие их яиц.
Переход к обитанию в почве сопровождался выработкой разнообразных физиолого-морфологических приспособлений у различных групп животных. Так, отсутствие света приводит к потере зрения и пигмента покровов у обитателей глубоких почвенных горизонтов. У животных, поселяющихся в верхних слоях почвы или часто выходящих на ее поверхность, пигментация и зрение сохраняются.
При переходе от водной среды к почвенной многие животные вырабатывают приспособления, защищающие их от высыхания в случае недостатка влаги в почве. Например, у полихет, немертин, турбеллярий отмечается утолщение наружных покровов и снижение их проницаемости. Некоторые полихеты имеют мощно развитые кутикулы, особенно выраженные на голове, где они образуют подобие головной капсулы. Кроме того, происходит развитие желез, выделяющих большое количество слизи.
Следует, однако, подчеркнуть, что кутикула, несмотря на снижение своей проницаемости, у почвенных форм продолжает оставаться пригодной для диффузии дыхательных газов и испарения, благодаря чему многие беспозвоночные почвы дышат всей поверхностью. Это оказывается возможным прежде всего в силу высокой насыщенности почвенного воздуха влагой.
В случае же обитания в воздушной среде требуется значительно более эффективная защита от высыхания, исключающая кожное дыхание (Гиляров, 1949, 1977). У членистоногих, приспособленных к жизни в сухой атмосфере, такая защита связана с образованием на их покровах тончайшего слоя эпикутикулы, состоящей из восковых и жироподобных веществ.
Появление таких водонепроницаемых покровов неизбежно требует развития специальных органов дыхания — трахей.
Отсутствие эпикутикулы у влаголюбивых обитателей почвы — мокриц, губоногих многоножек, камподий, ногохвосток, личинок насекомых и др. — позволяет им использовать кожное дыхание. Такое дыхание, однако, делает многих животных весьма зависимыми от влажности почвы и вынуждает их к миграциям в глубь почвы в более влажные горизонты, а также к вторичному использованию метаболической воды и др.
Потребность в насыщенном влагой воздухе у почвенных животных с кожным дыханием нередко определяет конкретные особенности их экологии. Так, пустынная мокрица, по Н. А. Димо (1945), в Средней Азии встречается на лёссовых почвах, где она укрывается в норах, уходящих на глубину до 1 м, что позволяет ей дышать воздухом, насыщенным влагой. На легких же почвах отсутствуют благоприятные условия для обитания мокриц, что объясняется прежде всего невозможностью рыть глубокие норы с маленьким диаметром в горизонтах с рыхлым сложением.
Приспособительные изменения отмечаются у почвенных животных и при оплодотворении (Криволуцкий, 1969). Установлено, что характер оплодотворения зависит от влажности среды. У водных животных обычно наблюдается наружное оплодотворение, при котором происходит выбрасывание половых продуктов в воду. У обитателей воздушной среды доминирует внутреннее оплодотворение (введение самцами спермы в половые протоки самки) — в противном случае в сухой атмосфере яйцеклетка и сперма быстро бы погибли. У почвенных же животных имеются разные варианты «наружно-внутреннего» оплодотворения, при котором в простейшем случае оставленная самцом на поверхности субстрата капелька спермы подбирается самкой половыми крышками.
К числу важных адаптаций следует отнести приспособленность многих животных почвы к перенесению сильных физических воздействий. Например, такие мелкие обитатели почвы, как тихоходки, обладают необыкновенной способностью быстро впадать в анабиоз и переносить облучение высокими дозами рентгеновских и ультрафиолетовых лучей, высокие концентрации кислот и др. Показано, что этих животных можно без вреда держать 20 мес при температуре —190° С. Такая поразительная выносливость тихоходок, сочетающаяся со способностью распространяться ветром на любые расстояния, позволяет им заселять практически всю поверхность суши. Это характерно и для многих других обитателей почвы — нематод, микроарт- ропод, простейших, устойчивость которых к неблагоприятным воздействиям факторов среды очень велика. Например, коллемболы — один из отрядов низших бескрылых насекомых (вторая по численности группа микроартропод) — поразительно устойчивы к низким температурам и нередко бывают активны да-
же в мерзлой почве, а развитие их яиц не прекращается вплоть, до 2—3° (Криволуцкий, 1969).
Отмечается также изменение характера движения при переходе к обитанию в почве. Например, полихета Lycastopsis ато- boitiensis при обитании под опавшими листьями передвигается,, как сколопендра, по поверхности субстрата, но в почве она движется, как земляной червь.
У многих почвообитающих животных, переходящих к жизни в почве, наблюдается изменение формы тела. Отмечено, что обитание в системе мелких полостей между почвенными частицами приводит к значительному'уменьшению размеров корненожек по сравнению с обитателями водной среды. Измельчению животных способствуют также недостаток пищи и низкое содержание Ог. Мелкие размеры клетки обеспечивают более быструю диффузию в нее кислорода.
Для корненожек, живущих в почве, характерно также отсутствие раковинок сложной формы, среди которых преобладают удлиненно-овальные, сферические или полусферические. Выступы у раковинок — иглы, шипы, имеющиеся у полуводных и водных форм и способствующие, по-видимому, фиксации на листьях водных растений и увеличению поверхности, — у почвенных видов развиты слабо или полностью отсутствуют (Гельцер, Кор- ганова, 1976; и др.).
Значительную роль в заселении корненожками почвы сыграло образование плоской вентральной поверхности (подошвы) и приобретение полусферической формы, позволившей организму лучше использовать промежутки между почвенными частицами неправильной конфигурации. Форма и величина раковинок в значительной мере определяют преимущественную приуроченность разных видов корненожек к разным горизонтам профиля. Так, сжатые дорсо-вентрально раковинные амебы в основном обитают в подстилке. Между твердыми частицами гумусового горизонта живут довольно крупные полусферические виды. В нижние горизонты с плотным сложением и малым количеством гумуса корненожки почти не проникают. Здесь лишь отмечены единичные экземпляры рода Plagiopyxis (Bonnet, 1964; Schonbom, 1973).
Значительные изменения формы и размеров тела в связи с обитанием в почве наблюдаются у панцирных клещей (Криволуцкий, 1977). Приспособление к обитанию в глубоких горизонтах у клещей заключается в тонких покровах, коротких ногах, удлиненной форме тела, отсутствии крупных щетинок. Поверхностно обитающие виды имеют иную форму.
Оценивая характерные эволюционные особенности почвообитающих животных в целом, в свете имеющихся исследований по данной проблеме можно отметить следующее.
Прежде всего животный мир почвы (по сравнению с наземным) более архаичен. Так, для широко распространенных низших почвенных беспозвоночных характерна сапрофагия — питание разлагающимися растительными и животными остатками. Растительноядность же (фитофагия) распространена среди почвенных беспозвоночных значительно реже, причем главным образом среди более высокоорганизованных форм (Гиляров, 1949, 1968).
Другими показателями архаичности животного населения почв являются наличие примитивных черт организации и замедленность эволюции у ряда его представителей. Замедленность эволюции почвообитающих животных по сравнению с наземными была рассмотрена Д. А. Криволуцким (1977) на примере панцирных клещей (орибатид) почвы. Например, роды Culto- ribula и Achipteria живут уже минимум 140 млн. лет, в то время как среднее время существования рода у лошадей — 5,6 млн. лет, хищных млекопитающих — 6,5 аммонитов — 20 и даже у пелиципод — этой медленно эволюционирующей группы — 78 млн. лет. Причина медленной эволюции почвенных ори- ‘батид автором связывается прежде всего со стабильностью среды их обитания, где постоянно сохраняется высокая влажность воздуха, сглажены колебания температуры, а инсоляция незначительна.
При переходе от наземного существования к обитанию в почве у некоторых видов наблюдается упрощение организации. Так, отмечается редукция защищающих от высыхания приспособлений у форм, переходящих от открытого к почвенному образу жизни, например у слизней (Гиляров, 1949).
Архаичность животного мира почвы в известной мере проявляется и в высокой степени постоянства ее населения по своему составу,в различных местностях (по сравнению с наземными организмами), что связано с более слабыми колебаниями влажности и температуры почвы. Это обнаруживается особенно отчетливо при группировке почвенного населения по крупным систематическим категориям.
Указанное своеобразие животного мира почв — следствие •особенностей почвы как среды обитания, главными чертами которой являются буферность (способность противостоять резким изменениям основных своих параметров) и значительное разнообразие почвенных условий в пределах профиля: каждый горизонт — особая экологическая ниша. Благодаря этому почвенные животные, с одной стороны, защищены от экстремальных воздействий факторов внешней среды, а с другой — они обладают возможностью выбора подходящих им условий в пределах профиля, что позволяет им избегать дальних и длительных миграций.
Данные свойства почвы как среды обитания сыграли исключительную роль в эволюции животного мира, одной из характерных особенностей которой явилось постепенное освоение не только почвы, но и других сред (табл. 50), в том числе воздушной, требовавшее, как правило, значительных адаптационных изменений организма. Тенденция к выходу на поверхность от-
мечена даже у таких типичных обитателей почвы, какими являются дождевые черви.
Таблица 50
Местообитания личинок 100 семейств жуков (Гиляров, 1949)
| Место обитания | Почва (включая Ао) | Мертвая и гнилая древесина, детрит и т. д. | Внутри живых растительных тканей | На поверхности листьев, открытые местообитания | На животных | В воде | Неиз вестно |
| Встречае мость | 30(+4) | 33(4-12) | 10(+9) | К+2) | 1(+1) | 8(+2) | 15 |
Эволюция в разных систематических группах этих червей идет по пути перехода от жизни в почве, где они питаются перегноем, к питанию на ее поверхности опадом, но при использовании почвы как среды обитания. Следующий этап — это полный переход к жизни вне минеральных слоев почвы: в лесной подстилке или в растительном войлоке, в гниющей древесине, под мхом на стволах деревьев. Выход на поверхность дождевых червей сопровождается серьезными морфофизиологическими преобразованиями.
Сказанное свидетельствует о том, что наземный образ жизни требует глубоких адаптационных изменений, которые были бы весьма трудно осуществимы, если бы имел место сразу резкий переход от водной среды к обитанию на поверхности земли без промежуточного звена — почвы. Нам представляется, что для ряда групп животных такой переход оказался бы невозможным или же потребовал бы гораздо большего времени, что привело бы к общему снижению темпов биологической эволюции и ее упрощению. Иными словами, без почвы оказалось бы невозможным то разнообразие наземных форм, которое имеет место в настоящее время.
Данному выводу не противоречит положение о замедленных темпах эволюции собственно почвенных форм, так как, во-первых, без почвы этих форм могло бы вообще не быть, а во-вторых, не все обитатели почвы эволюционируют медленно. Например, в группе почвенных клещей, характеризующейся в целом замедленным формообразованием, выделяются экологические группы, эволюция которых протекает более интенсивно (Криволуцкий, 1977). Так, если у обитателей глубоких слоев почвы формообразование действительно весьма замедленно, то у клещей, живущих в поверхностных горизонтах, оно намного интенсивнее. Более активно эволюционируют относительно слабозащищенные тонкопанцирные клещи, обитающие в наиболее динамичной части почвы — ее скважинах и трещинах.
Оценка темпов эволюции по числу эндемичных родов у разных экологических групп клещей показала, что больше всего эндемиков у обитателей почвенных скважин (табл.51).
Буферность и внутрипрофильное экологическое разнообразие почвы благоприятствуют успешному осуществлению ею и
Таблица 51
Количество эндемических родов клещей в разных почвенных экологических группах (Криволуцкий, 1977)
| Почвенные экологические группы | Число эндемиков | Процент эндемиков | Всего родов |
| Обитатели: глубоких горизонтов................................ | 3 | 30 | 10 |
| открытой поверхности............................ | 48 | 32 | 148 |
| мелких почвенных скважин .... | 15 | 41 | 37 |
| Неспециализированные формы .... | 5 | 21 | 24 |
| Всего . . . | 71 | 32 | 219 |
функции хранителя ряда древних форм, которая по-настоящему еще не изучена и не оценена, хотя и имеются факты, свидетельствующие о ее проявлении.
ЗАКЛЮЧЕНИЕ
Рассмотрение различных категорий функций почвы в системе ее биогеоценотических, ландшафтных и биосферных взаимодействий позволяет прийти к заключению, что экологическая многофункциональность почвы является ее фундаментальной, чертой. Однако до недавнего времени ей уделялось явно недостаточно внимания, что вело к общей крайне упрощенной и далеко не полной трактовке роли почвы в природных процессах и к недооценке необходимости сугубо дифференцированных приемов ее хозяйственного использования.
Выявление многочисленных функций почвы позволяет по- новому представить значимость ее в сохранении экологического благополучия планеты, более глубоко и всесторонне оценить сущность и разнообразие взаимодействий почвенной оболочки Земли с различными геосферами.
Вместе с тем становится очевидной важность дальнейшего, усиленного изучения закономерностей функционирования почвы как особой динамической природной системы. В ходе этого изучения возникает ряд взаимосвязанных задач и перекрещивающихся проблем. К их числу следует отнести прежде всего необходимость устранения диспропорции между исследованием относительно статичных параметров почвы и изучением ее мобильных составляющих, а также происходящих в ней процессов. Первые, как правило, изучаются чаще и в большем объеме, чем вторые.
В этой связи уместно привести весьма актуальные сегодня слова Б. Б. Полынова: «Даже самое поверхностное знакомство с историей развития наших представлений о земле, земной коре, слагающих ее горных породах и о составляющих эти последние минералах обнаруживает, что одним из крупных факторов этого развития является борьба между идеями двух различных категорий. Идея постоянства и родственные ей — покоя, равновесия и косности — боролись, в известной мере борются до настоящего времени с идеями текущего процесса, движения и превращения. Можно, по-видимому, утверждать, что прогрессивные эпохи этой истории связаны с победами второй категории» Ч
Другая, не менее злободневная, проблема — координация почвенно-генетических и экологических программ и расширение числа исследований, которые проводились бы на общих объектах по согласованным методикам на основе общей методологии. Почвенные системы — весьма сложные, многокомпонентные образования с высокой пространственно-временной изменчивостью, поэтому изучение законов их жизни и функционирования несмотря на свою притягательность, первостепенную теоретическую и практическую значимость — весьма трудоемкая задача, успешное решение которой предполагает тесный союз многих специалистов-единомышленников. Перед учеными-почвоведами стоит задача — своевременное усвоение почвенно-экологической информации, ее обобщение и синтез.