Функция стимулятора и ингибитора биохимических и других процессов
Данная функция почвы обусловлена тем, что в нее поступают разнообразные продукты метаболизма растений, микробов, животных — аминокислоты, белки, витамины, спирты и др., которые могут стимулировать или угнетать жизнедеятельность живых организмов (Троицкий, 1960; Пейве, 1961; Орлов, 1974; Дроздова, 1977; Галстян, 1978; и др.).
К настоящему времени накоплен значительный материал ио взаимному влиянию живых организмов посредством поступающих в почвенную и воздушную часть биогеоценоза метаболитов (рис. 5, табл. 15—17). По отношению к растениям и микроорганизмам эти данные обобщены в таких монографиях, как «Микроорганизмы почвы и высшие растения» Н. А. Красильникова, «Растительные выделения и их значение в жизни фитоценозов» В. П. Иванова, «Биохимические взаимовлияния древесных растений» М. В. Колесниченко и др. Имеются обобщающие работы и по взаимовлиянию животных посредством физиологически активных веществ, выделяемых во внешнюю среду, например «Телергоны — химические средства взаимодействия животных» Я. Д. Киршенблата.
Рассматриваемая проблема по отношению к почвенной экологии имеет несколько аспектов, среди которых к числу наиболее важных относится выявление вклада почвенных биохимических взаимовлияний в общую динамику экосистем и формирование биологической продукции. Практика показывает, что в ряде случаев существенные стороны жизни наземных биоценозов контролируются рассматриваемой функцией почв.
В качестве примера можно привести почвоутомление, когда почвы снижают свою производительную способность несмотря на достаточное количество в них элементов питания и благоприятные климатические условия. Обычно это происходит на пахотных землях при монокультуре. Почвоутомление может
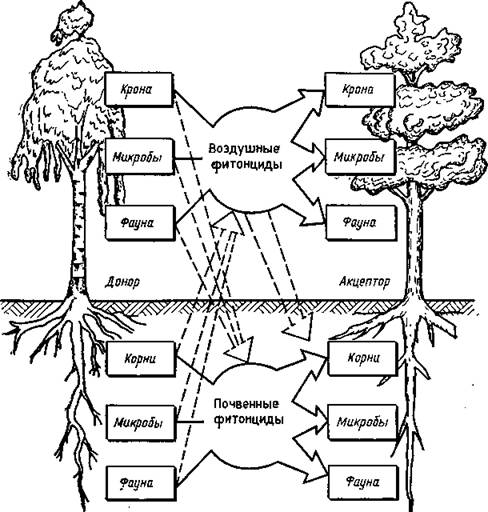
Рис. 5. Схема влияния фитонцидов одного растения на другое (Колесниченко, 1976)
быть вызвано развитием специфических патогенных микроорганизмов, паразитирующих на определенных видах растений, увеличением засоренности посевов сорняками и ухудшением водновоздушного режима почвы.
Нередко отмечается угнетение растений под действием корневых выделений. Явления самоугнете- ния отмечены у костра безостого, гваюлы и др. (Иванов, 1973). Угнетение одного вида другим неоднократно наблюдалось у лесных насаждений. Так, показано (табл. 18) отрицательное биохимическое влияние на дуб ясеня, вяза, осины, сосны (Колесниченко, 1976).Выделения определенных растений могут влиять на развитие других растительных организмов не только отрицательно, но и положительно. Так, при исследовании взаимоотношений древесных пород установлено положительное влияние на дуб выделений липы мелколистной и клена остролистного. Отмечено также положительное биохимическое взаимовлияние (табл. 19) сосны и лиственницы (Колесниченко, 1976).
Таблица 15
Биохимические влияния древесных пород (Колесниченко, 1976)
| Главная порода (акцептор) | Доноры фитонцидов | |
| активаторы | ингибиторы | |
| Дуб обыкновенный | гледичия обыкновенная, жи- | акация белая, береза бородав |
| молость татарская, клены остролистный, полевой, татарский, лещина обыкновенная, липа мелколистная, орех грец- | чатая, вязы обыкновенный И’ мелколистный, клен ясенелистный, осина, сосна обыкновенная, скумпия, тополь- канадский, ясени обыкновенный и пушистый акация желтая, береза боро | |
| Сосна обыкновенная | кий, свидина кроваво-красная лиственница сибирская, скум- | |
| Лиственница сибир | ПИЯ вяз обыкновенный, дуб лет | давчатая, дуб обыкновенный,, жимолость татарская береза бородавчатая |
| ская Береза бородавчатая | ний, клен остролистный, липа мелколистная, сосна обыкновенная, ясень обыкновенный клен остролистный, липа мел | вяз обыкновенный |
| Вяз мелколистный | колистная, лиственница сибирская, ясень зеленый ирга круглолистная, клен та | бузина красная, лох узколист |
| (перистоветвистый) | тарский, ясень пушистый, | ный, смородина золотистая,. |
| Тополь канадский | скумпия акация белая, акация желтая, | тополь канадский береза бородавчатая, вяз мел |
| (дельтовидный) | жимолость татарская, клен | колистный, бузина красная |
| Орех грецкий | татарский, ольха черная, скумпия, ясень пушистый | дуб обыкновенный |
Примечание.
В группу активаторов включены и породы индифферентные^Таблица 16
Отклонение скорости накопления Р32 в стволиках, % от нормы (Колесниченко, 1976)
| Растение-донор и его возраст, лет | Растение- активатор | Действие фитонцидов | ||
| воздушных | почвенных | воздушных и почвенных | ||
| 4 года | ||||
| Ясень обыкновенный....................... | дуб | не опр. | —29 | не опр. |
| Дуб черешчатый................................ | ясень | 0 | + 17 | +10 |
| Липа мелколистная........................... | дуб | +17 | +45 | +52 |
| 40 лет | ||||
| Липа мелколистная........................... | дуб | +29 | +122 | +юг |
| Вяз обыкновенный............................ | дуб | —80 | —113 | +109 +6S |
| Клен остролистный........................... | дуб | +20 | +40 | |
Таблица 17
| Доноры фитонцидов | Растение. акцептор | Фотосин тез | Накопление Р32 | Рост стволика | |
| по высоте | по диаметру | ||||
| Липа | дуб | +42 | +38 | +11 | +33 |
| Клен | дуб | +23 | +20 | +21 | +33 |
| Дуб | липа | +38 | +40 | +33 | +36 |
| Дуб J | клен | +36 | +188 | +29 | +60 |
Отклонение ряда показателей жизнедеятельности растений при воздействии почвенных фитонцидов, % от нормы
(Колесниченко, 1976)
Часто наблюдается и безразличное отношение растений как к собственным, так и к чужим корневым выделениям. Например, конопля, картофель, пшеница, ячмень, кукуруза не проявляют признаков самоотравления через корневые выделения (Иванов, 1973). Индифферентны к выделениям друг друга дуб и ель (Колесниченко, 1976).
Говоря о конкретных проявлениях почвенной функции стимулятора и ингибитора в связи с выделениями растений, необходимо отметить сложность установления ее конкретного результирующего действия. Это обусловлено несколькими причинами. Одна из них — возможность противоположно направленных влияний воздушных и почвенных выделений, которые, в частности, обнаружены у дуба и березы бородавчатой. Воз-
Отклонение интенсивности фотосинтеза, % от нормы при биохимическом взаимодействии дуба и других древесных пород
(Колесниченко, 1976)
Таблица 18
| Воздействующая древесная порода | Влияние на дуб фитонцидов | Влияние фитонцидов (воздушных и почвенных) дуба | |
| воздушных | воздушных и почвенных | ||
| Дуб летний4.................................................... | 0 | 0 | 0 |
| Ясень обыкновенный .................................. | —11 | —19 | —4 |
| Вяз обыкновенный........................................ | 0 | —13 | 0 |
| Липа мелколистная....................................... | -6 | +12 | 0 |
| Клены: | |||
| остролистный................................................. | +11 | +7 | —1 |
| полевой.......................................................... | —3 | 0 | |
| татарский....................................................... | +7 | +8 | 0 |
| Жимолость татарская.................................. | +12 | — | |
| Береза бородавчатая.................................... | +29 | —21 | +17 |
| Осина ............................................................ | —7 | —14 | +12 |
| Сосна обыкновенная ................................... | — | —12 | —5 |
| Ель обыкновенная........................................ | —4 | +1 | — |
Таблица 19
Отклонение интенсивности фотосинтеза, % от нормы . при биохимическом влиянии сосны и других древесных растений
(Колесниченко, 1976)
| Древесная порода | Влияние на сосну фитонцидов | Влияние фитонцидов сосны (воздушных и почвенных) | |
| воздушных | почвенных и воздушных* | ||
| Береза бородавчатая.................................... | —13 | —8 | +3 |
| Лиственница сибирская................................ | +12 | +16 | +5 |
| Дуб летний..................................................... | не оггр. | —5 | —12 |
| Вяз обыкновенный........................................ | » | —7 | не опр. |
| Жимолость татарская.................................. | —25 | —35 | » |
| Акация желтая................................... < . . | —7 | 0 , | » |
душные фитонциды березы Действуют положительно, а почвенные — отрицательно (см. табл. 18). Другой важный фактор, который необходимо постоянно учитывать, — зависимость действия выделений живых организмов от их концентрации. Так, малые концентрации выделений дуба оказывают положительное влияние на сосну, а большие — отрицательное. Аналогичная картина наблюдается и у сосны и березы (Колесниченко, 1976).
Таким образом, косвенная оценка рассматриваемой функции почв по отношению к конкретному растению только на основании учета особенностей видового состава биоценоза далеко не всегда возможна. Поэтому все большее значение приобретает прямое определение биологически активных соединений непосредственно самой почвы (Галстян, 1978).
Трудности диагностики активаторно-ингибиторных особенностей почв по отношению к тому или иному виду растения обусловлены также эффектами взаимодействия и изменчивости других биогеоценотических функций почвы. Так, лучший рост сосны с небольшой примесью березы, наблюдавшийся в опытах (Кабашникова, 1971), может быть связан не только с тем, что выделения березы в небольших концентрациях положительно влияют на сосну, но и с обогащением почвы листовым опа- дом березы, т. е. усилением почвенной функции источника элементов питания.
Рассматриваемая функция почвы также тесно зависит от выделений микроорганизмов, которые оказывают большое влияние на питание растений (Красильников, 1960). Существуют микробы-антагонисты, подавляющие рост чуждых им микроорганизмов путем выделения веществ типа антибиотиков.
В связи с тем что микроорганизмы образуют сообщества в ризосфере растений, суммарный результат (активация или ингибирование жизнедеятельности растений) будет зависеть не только от особенностей фитоценоза, но и видового состава микробоценоза.* Необходимо отметить, что состав и активность микроорганизмов ризосферы одного растения может изменяться при действии на них корневых выделений другого растения при совместном произрастании.
Говоря о механизмах биохимического взаимовлияния живых организмов в почве, следует отметить их многоплановость. Можно вычленить рассмотренное выше непосредственное действие самих продуктов выделения и опосредованное влияние метаболитов через их воздействие на доступность элементов питания и изменение pH почвы.
Примером влияния метаболитов на пищевой режим почвы может служить усвоение элементов питания из нерастворимых органических веществ под действием внеклеточных ферментов растений и микроорганизмов. Известно также, что корни растений выделяют целый ряд органических кислот (яблочную, щавелевую, янтарную и др.), с помощью которых происходит растворение и усвоение ряда минеральных соединений из почвы (Колесниченко, 1976).
Изменение pH почвенных растворов под действием выделений живых организмов установлено давно. Хорошо известно подкисляющее влияние корневых систем хвойных пород. Например, в зоне распространения корней сосны концентрация водородных ионов выше (на 0,2—0,4, а иногда 0,5—0,8), чем за ее пределами, (Ковда, 1973). Влияние на pH почвы оказывается
• Таблица 20
Требования различных растений к реакции среды (Авдонин, 1Э82)
| Растение | Оптимальный интервал pH | Растение | Оптимальный интервал pH |
| Рожь . . ............................. | 5,5—7,5 | Клевер................ «... | 6,0—7,0 |
| Овес................................... | 5,0—7,7 | Люпин................................ | 4,5—6,0 |
| Пшеница яровая .... | 6,0—7,5 | Тимофеевка...................... | 5,0-6,5 |
| Пшеница озимая .... | 6,3—7,6 | Костер............................... | 7,0—7,5 |
| Ячмень •............................ | 6,8—7,5 | Капуста............................. | 6,7—7,4 |
| Кукуруза........................... | 6,0—7,0 | Помидоры........................ | 6,3—6,7 |
| Горох................................. | 6,0—7,0 | Морковь............................ | 5,5—?,0 |
| Фасоль.............................. | 6,4-7,1 | Огурец............................... | 6,4—7,0 |
| Соя..................................... | 6,5—7,1 | Рис..................................... | 4,0—6,0 |
| Просо................................ | 5,5—7,5 | Чечевица........................... | 5,5—7,2 |
| Гречиха............................. | 4,7-7,5 | Вишня................................ | 5,7-6,5 |
| Свекла кормовая .... | 6,2—7,5 | Брюква.............................. | 4,8—5,5 |
| Картофель........................ | 5,0—5,5 | Сераделла......................... | 5,4-6,5 |
| Турнепс ............................ | 6,0—6,5 | Лисохвост......................... | 5,3—6,0 |
| Свекла сахарная .... | 7,0—7,5 | Райграс ............................. | 6,8—7,5 |
| Конопля ........................... | .7,1-7,4 | Редис ................................ | 5,5—7,3 |
| Лен..................................... | 5,9—6,5 | Салат................................. | 6,0—6,5 |
| Подсолнечник........................................... * | 6,0—6,8 | Лук..................................... | 6,4—7,9 |
| Люцерна............................ | 7,0—8,0 | Хлопчатник....................... | 5,5-7,3 |
| Чайный куст ..*.... | 4,5-6,0 |
важным фактором взаимодействия живых организмов, поскольку оптимальное развитие многих из них может осуществляться в достаточно узком диапазоне pH (табл. 20). Так, М. В. Марков на основании обобщения имеющихся материалов приводит следующую шкалу кислотности почвы, показывающую пределы pH, обеспечивающие наилучшие условия для роста и развития культурных растений: люпин хорошо растет при pH 4—5; картофель — при pH 5; овес, лен, рожь — при pH 5—6; клевер, горох, пшеница — при pH €—7; свекла — в узком пределе при pH около 7; люцерна — при pH 7—8; хлопчатник при' pH 7,5— 8,5. При рН8,5 — высокой щелочностью (Иванов, 1973).
Активаторно-ингибиторная функция тесно зависит не только от характера метаболитов живых организмов, поступающих в почву, но и от динамики других ее компонентов. Так, большое значение имеет изменчивость влажности почвы, существенно влияющей на динамику метаболитов, поступающих в нее. Установлено, что обмен корневыми выделениями происходит в широком диапазоне почвенной влажности — от 25 до 90% полной влагоемкости. Однако интенсивный обмен корневыми выделениями и их активное биохимическое взаимовлияние наблюдаются при влажности около 70%. Уменьшение или увеличение влажности почвы по сравнению с оптимальными значениями вызывает торможение поглотительной деятельности корневых систем и резкое снижение обмена корневыми выделениями.
Одним из практических следствий изучения изменчивости активаторно-ингибиторной почвенной функции является учет полученных данных для оптимизации структуры посевов. Обращается внимание на перспективность специальных смешанных посевов и посадок с повышенным коэффициентом использования почвенного плодородия благодаря подбору видов с положительным взаимовлиянием и учету изменчивости сезонных и суточных ритмов корневых выделений. В таких посевах вещества, выделяемые корнями одного вида, могут поглощаться корнями другого вида с противоположно направленным ритмом поглотительно-выделительной деятельности и тем-самым препятствовать вымыванию из почвы соединений, поступающих из корневых систем. Эти соединения могут служить дополнительным источником элементов питания и выполнять роль активатора биохимических процессов в почве.
М. В. Колесниченко (1976) приводит факты снижения продуктивности чистых культур ели и сосны в Европе при выращивании 2—3 поколений в течение 200—300 лет. В лесах нашего Севера за 2—3 поколения чистых ельников бонитет их упал со II—III до IV—V классе. Одна из причин данного явления, по-видимому, почвоутомление в одновидовых лесах в связи с неблагоприятным биохимическим взаимовлиянием растений. В качестве одной из мер М. В. Колесниченко предлагает специально подобранные смешанные посадки, консортивные связи и более богатый видовой состав которых препятствуют развитию почвоутомления.
Учет динамики активаторно-ингибиторной функции почв нередко позволяет более объективно оценить причины происходящих изменений в биоценозе, поскольку они контролируются не только конкурентными отношениями видов, но и их биохимическими взаимовлияниями через посредство поступающих в почву продуктов метаболизма (Колесниченко, 1976). Эти взаимовлияния касаются не только растений и микроорганизмов, о чем говорилось выше, но и почвообитающих животных.
Выделяемые в среду, в том числе в почву и на почву, продукты внешней секреции обладают различным действием. Так, у очень многих видов насекомых половозрелые самки выделяют специфические пахучие вещества (одмихнионы), привлекающие самцов, которые могут воспринимать их на большом расстоянии, даже через толстый слой почвы.
Пахучие вещества наносятся животными на почву и служат им в качестве ориентиров при добывании пищи, а также могут выполнять роль меток, указывающих на занятость территории. Особое значение эти вещества имеют у общественных насекомых, которые оставляют пахучие следы от гнезда к источникам пищи. Большинство муравьев оставляет пахучие следы вдоль муравьиных дорог, нанося специфические вещества на поверхность почвы путем периодического прикосновения к ней брюшком или кончиком жала.
Свойства среды, в том числе и почвы, могут заметно влиять на способы выделения пахучих веществ и их количество. Своеобразно, например, выделение одмихнионов у некоторых му» равьев, живущих в пустынях и полупустынях. В связи с тем что по^ва и песок в местах их обитания сильно нагреваются солнцем, что мешает прикосновению брюшка к поверхности, эти муравьи выделяют одмихнионы прямо в окружающий воздух, быстро перебегая на длинных ногах по горячему песку, приподняв брюшко кверху (Киршенблат, 1974).
Многие животные могут выделять вещества, вызывающие у особей того же вида реакции тревоги, бегства, активной обо» роны. Вырабатываются и вещества (аминоны), выполняющие роль химических средств защиты от врагов, благодаря отталкивающему запаху, раздражающим и ядовитым свойствам.
Перечисленные действия выделений животных организмов во внешнюю среду существенно расширяют конкретные проявления характеризуемой почвенной функции активатора и ингибитора биохимических и некоторых других процессов, происходящих в почве или на ее поверхности.