Действие биологически активных агентов в сверхнизких дозах на биохимические и физиологические процессы
Мы начнем с цитаты из обзора, опубликованного одной из наиболее активных исследовательских групп, работающих в этой области: «В литературе можно найти огромное, постоянно растущее количество
доказательств чрезвычайно высокой чувствительности биологических объектов к химическим и физическим эндогенным воздействиям*.
Биологические объекты способны реагировать на присутствие пептидов, гормонов и ядов в концентрациях ниже 10”12 М. Чувствительные к запаху клетки реагируют на отдельные молекулы пахучих веществ. Зрительные рецепторы реагируют на отдельные кванты света.
Само по себе действие агентов, энергия которых на много порядков ниже тепловой, не удивительно. Как было подчеркнуто в первых главах этой книги, в случае кинетически неравновесных систем слабые силы могут вызвать сильный эффект. Мы не удивляемся, когда слабое нажатие пальцем на спусковой крючок ружья приводит к большим последствиям. Большинство биологических объектов и их компонентов принадлежат к кинетически неравновесным системам. Описание их поведения в рамках механики часто более адекватно, чем в рамках термодинамики.
Действительно удивительной характеристикой эффектов сверхмалых доз является немонотонность кривых в координатах доза-эффект. Авторы [81] называют это сложным полимодальным характером дозо- вой зависимости.
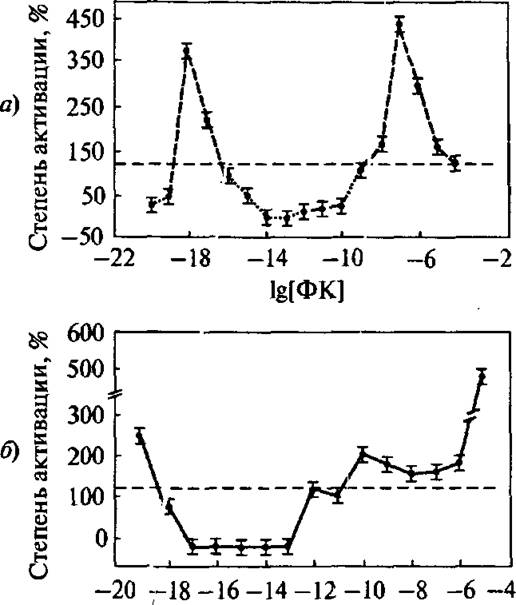
В качестве иллюстрации на рис. 5.1 приведены данные из работы Надежды Пальминой и др. [82]. Они исследовали воздействия различных биологически активных соединений (БАС) на активность фермента протеинкиназа С. На рис. 5.1 ясно видно существование двух областей максимальной ингибиторной активности при концентрациях 10“17 М и 10“12 М (для нормальных клеток). При концентрациях ингибитора 10“|0-10“14 М (для нормальных клеток) и 10”13- 10”17 М (для опухолевых клеток) можно видеть широкие области с практически нулевой активностью.
Было опубликовано множество аналогичных результатов по аномальному действию БАС в сверхнизких концентрациях на биохимические и физиологические процессы [84-88]. В этих случаях следует рассматривать действие отдельных молекул на мишень (клетка, связанный с мембраной рецептор, глобула белка и т. п.).Я не могу обсуждать здесь отношение этих эффектов, достоверность которых вне сомнения, к гомеопатии.
ІВІФК]
Рис. 5.1. Типичная кривая «доза—эффект». Зависимость активности протоин- киназы С от концентрации фенозана (активатор): а) нормальные клетки; б) опухолевые клетки
На самом деле возникают только два главных вопроса.
Первый сформулирован авторами в обзоре [81]. «Трудно понять даже саму возможность встречи отдельных молекул БАС с клеткой за разумный период втрёмени. Казалось бы, эти индивидуальные молекулы должны хаотичеок и блуждать в межклеточном пространстве перед контактом с мишенью. И только после сосредоточения их достаточного количества в межклеточном пространстве и перехода к равновесию система может реагирошть».
Авторы рассматривают ситуации, возникающие в двух предельных случаях: высокая и оче нь низкая концентрация БАС.
Молекула. БАС должна контактировать с рецептором на поверхности клетктл Учитывая разумные значения размера клетки (2-10 микрон) и коэффициента диффузии БАС (D — 10"3 см2/с), а также
возможность латеральной диффузии молекул БАС по поверхности мембраны [83], авторы г/риходят к выводу, что достаточно рецепции одной-десяти молекул БАС. Время рецепции (50-90) % растворенных БАС не превышают нескольких минут.
Если число молекул очень мало, нельзя предполагать существование концентрационного градиента.
Проведенные автором расчеты в рамках схемы Бернулли «попадание-промах» показали, однако, что дл я концентрации 10”17 М число молекул БАС в реакционном пространстве (0,5 мл) будет достаточно дая регистрации эффекта в течение времени эксперимента.
Второй вопрос касается причины полиэкстремального характера зависимости кинетических параметров исследуемого процесса от концентрации БАС. Авторы подчеркивают, что в случае сверхмалых доз сама концепция концентрации теряет свою ясность. Распределение частиц в реакционном объеме становится дискретным, и сильнее проявляется неоднородность распределении БАС в непосредственной близости от мишени. Это неизбежно приво дит к отсутствию воспроизводимости.
Бурлакова и ее соавторы [81] предложил^ остроумную модель, способную превратить локальные флуктуации концентраций БАС вблизи мишеней, мешающих регистрации эффекта* в механизм, генерирующий эффект. Для этого было сделано несколько предположений. Первое заключается в том, что каждый акт рецепции может вызвать два типа ответа клетки: «+» и «-», существенно превосходящие некий порог (это и есть второе предположение). Эти два типа ответа могут быть реализованы, если мишень может существовать в двух состояниях, например, с двумя возможными конформациями рецептора.
Предположив также несколько других характеристик мишени и механи зм их взаимодействия с БАС, авторы сформировали простые и в то же время общие схемы, объясняющие действие БАС в области сверхмалых доз.
Рассмотрим теперь другую простую модель, открывающую путь к пониманию эффектов, наблюдаемых при сверхнизких ко?нцентрациях БАС, действующих на клетки и другие мишени [89].
Экспериментальные результаты позволяют предположить, что при достаточно низких концентрациях БАС возникает некий параметрический резонанс, определяемый внутриклеточными процессами, вызванными БАС, и характеристическим временем поступления БАС (или продуктов их химического взаимодействия с компонентами клетки) к специфическим сайтам внутри клетки (мембранные рецепторы, активные центры ферментов).
Разумно предположить, что при сверхмалых концентрациях БАС лимитирующей стадией всего процесса будет диффузия молекул БАС из объема к поверхности клетки. Все другие стадии, например, химическая трансформация БАС, их связывание с активным или аллостерическим центром фермента и т.
п. происходят намного быстрее. Характерное время может быть таким образом определено как обратное от числа (Z) столкновений БАС с клеткой за единицу времени. Значение Z можно легко рассчитать с помощью уравнения Смолуховского:Z = 4xDRn. (5.1)
Здесь D — коэффициент диффузии молекул БАС, R — радиус клетки а п — число молекул БАС в единице объема.
Учитывая сказанное в главе 4 этой книги, представляется разумным выбрать в качестве характеристического параметра времени внутримолекулярного процесса, ответственного за наблюдаемый эффект, время конформационной релаксации определенных макромолекуляр- ных структур. Такими структурами могут быть: ферменты, регуляторы биосинтеза, рецепторы, связанные с мембранами, и т. п., локальные возмущения которых приводят к появлению неравновесного состояния. В целях упрощения мы в дальнейшем будем называть такие структуры ферментами.
Конкретный механизм суммарного процесса в данном случае не важен. Достаточно предположить, что внешний агент (например, БАС), поступая в клетку со средней частотой Z, определяемой по формуле 5.1, приводит к возникновению конформационно неравновесного
На самом деле в уравнении должна быть сумма радиусов сталкивающихся частиц. В нашем случае радиусом второй частицы можно пренебречь.
состояния фермента. Это состояние затем релаксирует с характеристическим временем г к новому состоянию, равновесному для фермента с присоединенным агентом. На определенной стадии релаксации активность фермента максимальна.
Для очень низких концентраций БАС, когда Z~x > т, почти все молекулы фермента находятся в начальном, равновесном состоянии с низкой активностью. При высоких концентрациях БАС, когда Z~x < т, почти все молекулы фермента будут в конечном равновесном состоянии с низкой активностью. Максимальную концентрацию неравновесных промежуточных состояний фермента с повышенной активностью можно ожидать при грубом совпадении значений Я-1 и т.
Не представляет труда рассчитать молярные концентрации БАС для мишеней с различными значениями R, соответствующие разным величинам Z~x — т (табл.
5.1). Был выбран коэффициент диффузии молекулы сахарозы в водном растворе (5Ю“10 м2с-1). Из 5.1 получаем:с«3-10-13т_1Д-’. (5.2)
Здесь с — молярная концентрация БАС, т — время релаксации (с), Л __ радиус мишени.
Поскольку имеют смысл только расчеты с точностью до порядка величины, можно опустить множитель 3, четыре выбранные размера покрывают мишени от макромолекул до больших клеток.
Легко видеть, что для мишеней с размерами клеток (1 мкм), можно ожидать появления экстремумов на кривых. Концентрация БАС — эффект в интервале 10”11 -10“15 М.
Естественно, приведенные здесь объяснения могут служить только указаниями на вероятность адекватности предложенных подходов. Исследования аллостерических ферментов при низких концентрациях эффектора и с параллельными измерениями времени конформационной релаксации белка после присоединения эффектора могут привести к экспериментальному подтверждению концепции.
Во всяком случае, ясно, что проблема странного поведения БАС при сверхнизких концентрациях может быть решена в рамках современной биофизики.
Таблица 5.1
Молярные концентрации (- Igc), соответствующие максимальной активности для различных значений R и т
| R, мкм^. | 10“2 | 10’1 | 1 | 10 | 100 |
| 10"2 | 9 | 10 | И | 12 | 13 |
| 0,1 | 10 | 11 | 12 | 13 | 14 |
| 1 | 11 | 12 | 13 | 14 | 15 |
| 10 | 12 | 13 | 14 | 15 | 16 |
Я не буду обсуждать здесь сенсационные эксперименты Бенвени- сты и corp. [90], посвященных активности некоторых БАС в растворах при концентрациях ниже 10~24 М.